
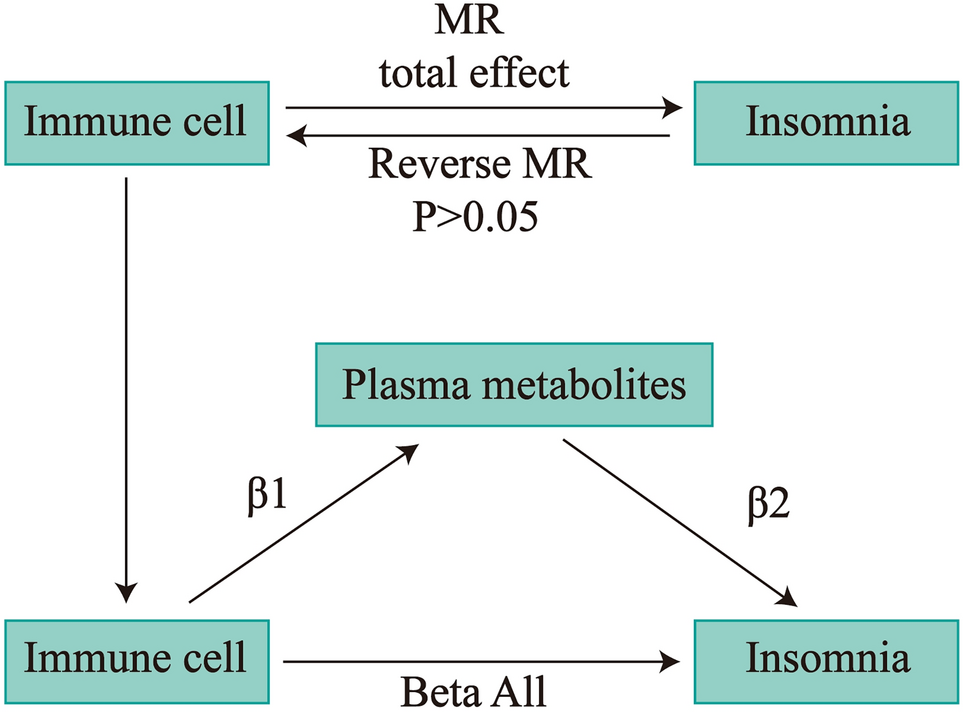
Remember me
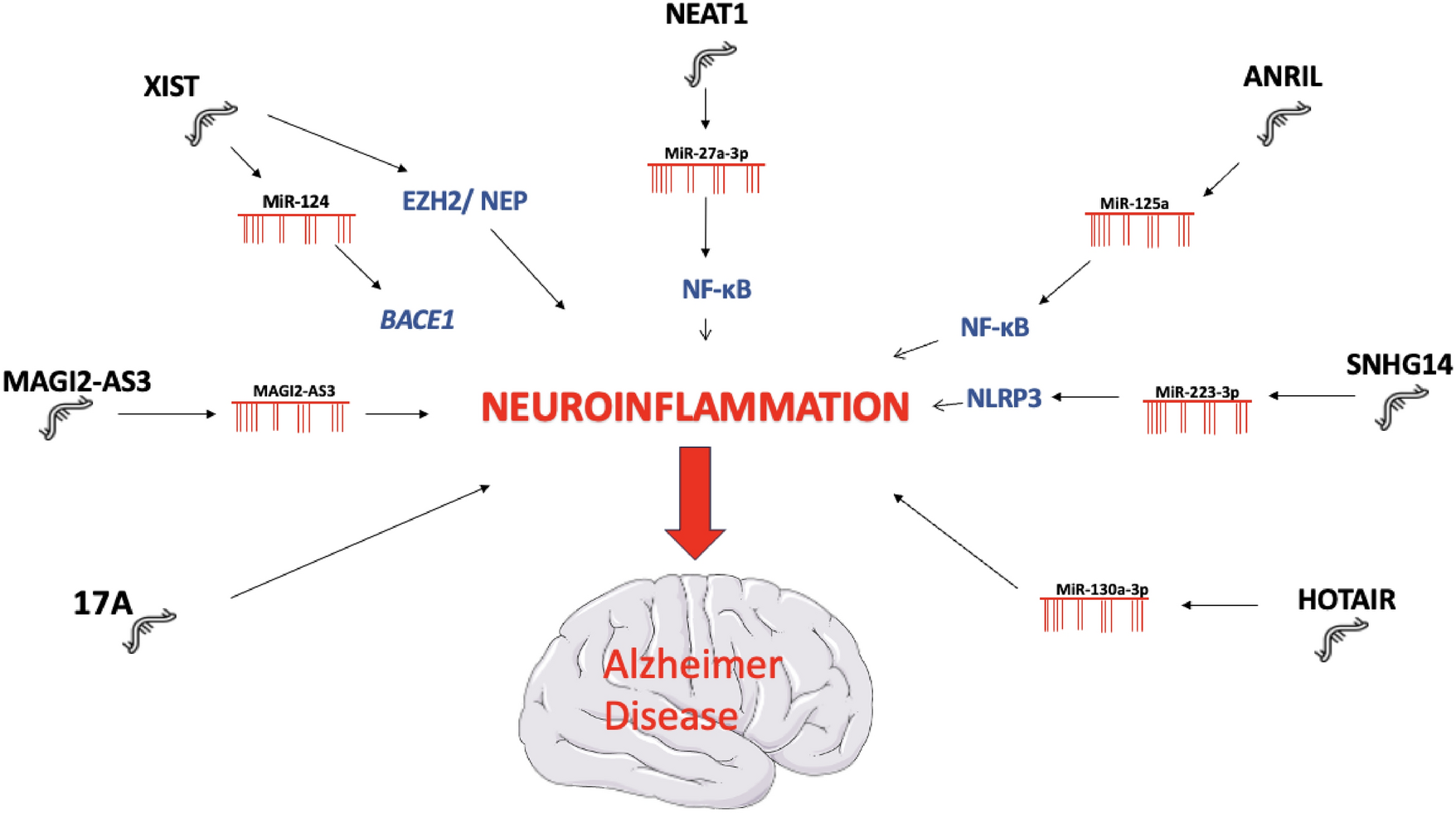
Microglia cells have a unique position in the immune response in the post-BBB interstitium as they are usually the first motile responders during perturbations within the extracellular biological matrix. This cell is mobile and highly amorphous and as such its characteristics can potentially be visually read by evaluating its shape and the sum of its three main structural components: the soma, the nucleus, and the dendrites. Much investigation has gone into studying its behavior and signaling during activation (Fernández-Arjona et al. 2017; Filipo et al. 2019; Kamphuis et al. 2016; Minami et al. 2012), this is the first reported evaluation within a cultured organotypic tissue model that activated a subset of MG within the tissue model with a toxic injury but left the rest of the MG in its inactivated baseline state. Thus, one of the enticing aspects of observing MG embedded within a viable tissue matrix is observing MG response events in-situ, by modifying (via toxicity) the immediate interstitium (intercellular space) amid a viable biologically active sample (as demonstrated in electrophysiology studies; Stanton et al., 1984). It is also important to note that maintains the microscopic structural layout of a mammalian brain is preserved in this tissue slice. The benefit of this condition is that it allows for evaluation of a spectrum of MG morphological changes that occur from being in a baseline state to that of being fully activated within the tissue context. However, prior to delving into these deep queries on how MG (and CNS cells) ascribes a genetic commitment that leads to stress related biochemical, proteomic changes, a reproducible clear cellular response must first be ascertained, in this study, such cellular response is morphology, and thus, how MG cell rearranges its cell membrane. Such is that this investigation into microglia’s morphology after precise toxic insult elucidates a nuanced cellular response, and in turn also revealed many details of MGs’ structural variation within its nascent CNS surroundings.
The first two characteristics, nucleus and cell body of the MG are intimately related as they are both stark visual landmarks whether MG is in its baseline state or its activated state. Albeit there are appreciable changes in size of MG when activated versus in baseline, the most consistent quantifiable change is the space between the nucleus membrane and the cell membrane, as mentioned in Graphs in Figs. 4 and 5). This is an important marker not just of size, but more so the rearrangement of MGs cell membrane borders. The amorphous nature of MG is one of its important characteristics and these stark changes to cell structure enable functions as it relates to MG’s baseline state or activated state. Thus, when MG is in its baseline state the cell membrane’s distance (the wider of the lateral 2 distances) from the nucleus is around 1 micron thick, thus it is presupposed that the rest of the membrane is thinly spread out covering a designated surface area as it conducts its baseline activities. Conversely when MG is activated the nucleus’ membrane distance from cell membrane is greater than 3 microns on average (up to 6 microns individually), which is a substantial change. This quantitative change can serve as an indicator of level of activation of MG. As such this distance can serve as a point of distinction by characterizing intermediate forms of MG, along with other ancillary visual characteristics: dendrites size, shape and nucleus placement. Irrespective of the two distinct mechanisms of cell death and injury, MG morphologically behaved in the same way at least in this rudimentary qualitative assessment of MG shape. These 2 models of toxicological insult (mercury and cyanide combined with 2-deoxy-glucose) leveraged the knowledge of toxicological agents’ mechanism of toxicity to induce a cellular response, activating MG from an M2 to an M1 state.
Understanding the meaning/functionality MG dendrites is paramount in the understating of MG’s inherent biology and response to toxic insult. One of the most defining characteristics of MG is the presentation of their dendrites (Fernández-Arjona et al. 2017; Masuch et al. 2016). As described in the results, there are usually 3 major dendrites and one of them bifurcates into 2–3 branches of similar width, which resembles a claw. This dendrite is usually off-center and, therefore, lateral, which indicates that these cells are asymmetrical. This asymmetry can be useful in determining the orientation of the cell and thus have a greater appreciation of its layout within the tissue matrix. Furthermore, the other dendrites serve some sort of functional weight, potentially distinguishing the functions of all these variety of dendrites and providing important information about these structural variations. These structural variations can offer information about how these dendrites are protruding and assist in several different functions, for example: the adhesion or anchoring to the extracellular matrix, reach out to neighboring MG to maintain adequate spacing (since they seldomly overlap), probe the extracellular fluid (for either signaling or indications of cell damage), or directly probe neurons, astrocytes (and other CNS cells), cellular address probes, etc. and other undiscovered functions. Conversely these dendrites involute during MG’s most activated states, however often dendritic stumps remain, what role do these dendritic remnants emanating from the ameboid MG cell body play when MG is conducting its activation duties? Taken together, the nature of the MG cell is one that is highly animated and has several different opportunities for scientific endeavors on how these cells can serve as biomarkers of extracellular adequacy. The morphological plasticity of MG is intrinsically connected to its response to toxic injury. The main tenet of this study was to examine a controlled signal to noise of MG according to its morphology presentation within a sample that had simultaneously healthy and injured tissue, such is why the toxicity was localized to only injure a portion of the hippocampus tissue sample.
Prinz et al. (2019) describe that establishing microglial states can be essential to their roles in homeostasis and disease, such example was postulated as the disease associated MG (DAM) that was genetically characterized by Keren-Shaul et al. (2017). This study ascribes the morphological state as a determinant of its behavior, ergo “structural variation”. Structural variation alludes to the benefit of a cell’s structure to a role it is executing in response to its surroundings. Structure is an important demarcation of cellular activity because of the cells biochemical, metabolic, proteomic, genetic, and adaptive commitment of maintaining a specific macro shape. Many of the descriptions of baseline microglia in the literature default this structural motif to that of it being in a state of “sensing” the extracellular environment. Taking into account the known roles of MG it is understood that this state is within its purview of its characteristic activities, a sentinel of extracellular medium’s adequacy. In a baseline state they are spread out evenly throughout the hippocampus tissue matrix and seldom overlap; this fine-stellate form is suggestive of a behavior such as sensing and probing the interstitium’ s integrity as well as the somatic cells they mount as they migrate through an extracellular region. Baseline MG size ranges from 30 to 40 microns, and the filaments that emanate from the cell body that is just 1 micron thicker than the nucleus compartment, and the filaments themselves are barely thicker than a micron in width. This is logical considering the generally accepted phenotypic job description of dendritic cells, it is virtually a cellular detector of extracellular conditions. Upon its activation, MG changes into a much different shape and therefore it is no longer optimized for detection, it metamorphizes into a compact globular circular shape in its most extreme form of activation. The activated shape is likely more amenable for endocytosis of cellular debris that occurs during injury, accelerated motility and/or cordoning off a space withing the interstitium as this enlarged shape decreases the space available through micro-openings in the interstitium from which motile cells lay their trails, or interstitial fluids flow. Spatial obstruction by activated MG could decrease the flow of non-optimal extracellular fluid contaminated with potentially harmful intracellular contents from cells with compromised membranes.
It is also important to note that MG activation does not seem to be a binary outcome, that is, there are other intermediate forms that can arise and participate in the protective, recovery or pathologic process. The most compelling piece of evidence of this is simply the appearance of these intermediate forms near regions of cellular stress. This region is found right outside the most central part of the spot, where all cells are presumed dead (Fig. 2B). Further, these changes can be characterized by membrane rearrangement, the appearance, and morphologies of dendrites and nucleus placement within the MG. MG morphology had been a binary outcome only baseline and activated forms would have been found in these areas. Therefore, considering that MG activation fall into some sort of spectrum, these forms can signify the state of the immediate matrix they are surrounded by, and thus have a better understanding of the initial stages of cellular perturbation as recognized by this primary sentinel immune cell’s morphological presentation.
As described in the results section (Graphs on Figs. 4, 5), one of the most quantitative ways to assign this morphological spectrum is to measure the distance of the nucleus membrane to the cell membrane as that underlines the extensive amount of membrane rearrangement MG undergo that happened after the cell polarizes. In between these 2 extremes, 5 different morphologies were identified using cues from the size and shape of the soma and dendrites as well as the overall length of MG (Fig. 7, Table 1). These are postulated to be stable morphologies since they are present even 2 h after the initial toxicological insult, and since there is no migration of MG observed, these are not interloper MG cells that migrated from other region (or from nearby blood vessel). There had been migration which it could have been postulated that these were migrant MG from other areas that were just arriving at the sites of injury and thus in the middle of the M2 to M1 metamorphosis. Therefore, these intermediate morphologies can be ascribed to roles that have not yet been elucidated.
Fig. 7
Spectrum of microglia’s 5 distinct structural variations (SV) in between baseline (M2) and activated (M1) state. In between the 2 known states of microglia M2 (baseline) and M1 (activated), 5 intermediate structural variations can be identified based on the shape of the soma, the contraction of the dendrites and the cells’ overall size. Structural variations greater than 3 indicate that microglia ceases being a fine-stellate form and is becoming a dense cellular particle. Bar = 5 microns
Table 1 Structural details about microglia's 5 different Structural Variation s between its baseline morphology (M2 state) and activated morphology (M1 state).
Collating the visual empirical data gathered in this study together, that is MG overall size, shape/size of soma and appearance of dendrites a pattern of other structural variation intermediates emerges. Namely 5 different intermediate structural variations become apparent (Fig. 7). These qualitative categorizations are subjected to the appearance of the soma, nucleus, and dendrites in a 2-dimentional plane that captures all three. The 2 extremes of structural variations are M1 (activated MG) and M2 (baseline MG); the former is a dense cellular particle that is globular and has no dendritic processes, the latter is the appearance of a nucleus enveloped by a thin cell body with several thin dendrites emanating out of it. Since based on the quantitative assessment, the membrane of the soma gains greater distance from the membrane of the nucleus, a qualitative assessment as to the orientation of the membrane is applied; that is, when and where this separation starts to happen. Also, how this qualitative characterization change in membranes’ distance relates to the thickness and length of the dendrites is appraised. In between the two extremes five morphological states are described:
Structural variation 1- The first indication of morphological changes after the baselines state is the widening of the soma as the space between the nucleus membrane and the cell membrane start to gain some distance, this distance improves the visualization of MG as membrane stain is more intense.
Structural variation 2- Is demarcated after the first separation of the nucleus and cell membranes, the soma begins to polarize, that is, the slightly thicker membrane begins to accrete at opposite ends which results in an uneven encirclement of the nucleus which renders the soma more oval than circular in shape; in addition, the soma is elongated to the point that the nucleus can be regionalized within, usually there is 10 microns (or 2 nucleus lengths) between the 2 polar ends of the soma. After the polarization/elongation, the soma begins to widen and the measurement of the lateral distance of the soma’s membrane to the nucleus’ membrane could be objectively measured (as was demonstrated in Graphs in Figs. 6, 7).
Structural variation 3- Is the further elongation of the soma to the point that it reaches 3 nucleus lengths (nucleus was consistently found to be 5 microns in this study), but its width starts to be greater than 7–8 microns wide and thereby lateral separation from nucleus membrane begins; another very important distinction is the widening and shortening of the dendrites. An important structural consideration to structural variation 3 is that the accretion of soma is beginning to yield a denser cellular particle, which means it is becoming a more obstructive presence within the space it occupies, an important spatial consideration. The immediate consequence of this size and shape change is the alteration of interstitial fluid flow and the passage of motile cells.
Structural variation 4- After max elongation is reached (usually 15–20 microns) presents as a highly visible MG that has a much bigger and thickened soma but still stellate as all the dendrites are also thickened. Notably its dendrites are shortening into the soma, also, the nucleus membrane will gain greater distance from the cell membrane.
Structural variation 5- the last step before full activation, the soma is even more globular, rotund, but most notably there are fewer dendrites and the ones left over appear as stumps, compared to how they looked previously. This metamorphosis does appear to bear some impact on the conditions of the unique interstitium of the CNS, namely in the redistribution of the area of this cell’s soma and the impact of this on the availability of interstitial space, which will be discussed at greater length in an upcoming manuscript related to intercellular space (Trejos, in preparation).
Taken together, breaking apart the main characteristics of MG (soma, dendrites, nucleus), their intermediate forms suggest that they could be objectively discerned as having distinct visually definable structural variations; this is important because they can be associated with states of cellular stress prior to cell death as many of these intermediate shapes appear within the injury region of the spot tox adjacent to the epicenter.
Furthermore, and importantly, although the measurements in this research were done manually (Figs. 4, 5), this quantification can illustrate a great potential image analysis tool to efficiently evaluate the relative activation of MG in and around injured tissue. This can be potentially applied to AI program algorithms that can detect and evaluate MG in greater detail, so long as the setting can be programmed to detect and discern the nucleus membrane to that of the cell membrane; the relative distance between these 2 walls can be easily measured.
Microglia characterization within the tissue suggest a cell that undergoes dynamic change and structurally acquiesces to its extracellular environment, likely why MG evacuating the tissue matrix exhibit stark differencesin shape from MG embedded within tissue (Fig. 2F, G). Utilizing this structural variation criterion can inform the relative response of a MG with regard to its response to a cellular event that indicates cell damage is occurring. In Fig. 7, the higher gradations of structural variations surround the activated state whereas conversely the lower gradations surround the baseline state. Having this response layout can add resolution to the level of cell injury and how the structural variations of MG can be utilized as a proxy for perturbations in CNS cellular or interstitial homeostasis. The offered resolution to cellular metamorphosis of MG should serve as important distinction points of its activity as it relates to its morphology that can suggest an anomalous interstitial condition (which arises from an injured cell that is about to lose viability) ergo a precursor pathological event that eventually leads to some CNS pathology (in this case pathology that arises from chemical toxicity). Importantly, this detectable/measurable precursor event, microglial metamorphosis, may offer better opportunity for understanding how dendritic/MG cells respond to post-BBB CNS cell stress; that is, if we confide in the ability of these cells to detect CNS cell injury prior to than the current most sensitive determinants (intracellular enzyme levels increases in extracellular fluids which suggests their leakage from damaged cells). These cellular events are likely antecedent to mass enzyme leakage which represent cell membrane rupture.
In conclusion, the microglia is a unique cell within the most exclusive extracellular matrix in the body, its 3 main characteristics, that is, size, dendrites, and cell body shapes allow the assignment of distinct morphological structures. Affirmations of cell stress that is biochemically verifiable and confirmations of MG shape shifting characteristics could most effectively be done in this culture model; further and probably most importantly, the initial events of cellular stress within the context of cellular architectural tissue matrix are isolated. Downstream biochemistries can be studied that ascribe certain genetic (as demonstrated by Keren-Shaul et al. 2017) or proteomic changes that can relate to MG’s morphology can be further characterized and studies in greater detail in this isolated sample without the noise presented by extra-matrix influences of blood-derived cells and chemokine signaling outside the interstitium; and conversely, much more contextually than in cell suspension culture isolation. And, albeit MG appears as an amorphous construct, it follows a certain structure pattern as it metamorphosizes, and thus, leads to 5 distinct intermediate definable structural variations between baseline (M2) and activated (M1) forms. Furthermore, detailed, and definable aspects of its cell body structure (and quantitative changes in membrane distances that can be amenable to algorithmic evaluation), nucleus placement, and number of dendrites indicate that perhaps MG’s shape is not as vague as it appears. Also, it is important to note that as MG reach the M1 state (structural variations 3 +), they become a denser cellular particle that can impact interstitial spacing and thus glymphatics (Nedergaard and Goldman 2020; Trejos et al., in preparation) and thereby interrupt interstitial flow. Such is that the data generated from this isolation indicated that the hippocampal tissue slice is a relevant biological quantum of contextualized cells that maintains initial immune cell activity, confirmable survival, and spacing profile of CNS tissue matrix after excision. The biochemical confirmation of a cellular stress and death using MTT along with PI and then evaluating the individual properties of the MG cell response as a dendritic cell represents a fortified rational biological sequence of events that can be isolated and explored further. This isolation can shed some further depth of biochemical, proteomic, and genetic understanding as to what happens when tissue is injured and how it concerts with immune cells’ morphological response. Nevertheless, the structural resolution offered describes how microglia behave morphologically (hereby presented as structural variations) and can be used to potentially attribute events antecedent to cell death within the post-BBB CNS and adds greater depth of knowledge to microglia’s role in this highly exclusive parenchymal compartment.
Comments (0)