
Remember me
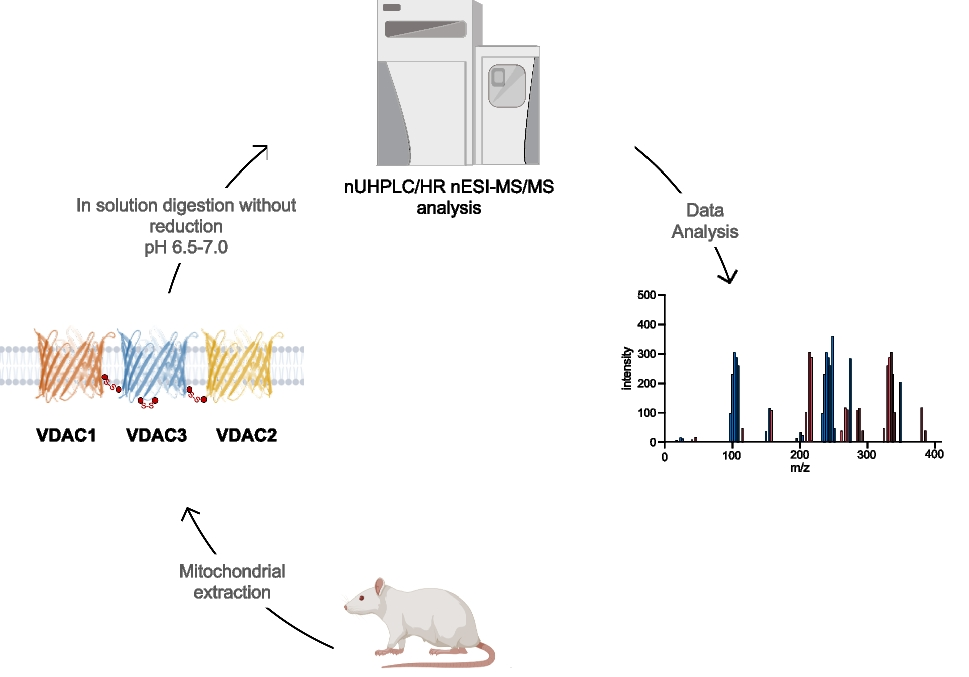

The sequence of rVDAC3 includes seven cysteines in positions 2, 8, 36, 65, 122, 165, and 229 (Fig. 1). The numbering adopted starts from Met1, which, although it is reported in the SwissProt database (Acc. N. Q9R1Z0), is absent in the mature protein [23]. Following post-translational modification, which eliminates the starting methionine, the cysteine encoded as the second amino acid becomes the N-terminal residue. The presence of a cysteine at the beginning of the N-terminal alpha helix is a unique feature of the VDAC3 isoform.
Fig. 1
Lateral and top view of rVDAC3. The structure of rVDAC3 is predicted by homology modelling, using mVDAC1 structure (pdb: 3EMN) as a template. Graphical representation was obtained by using PyMOL 1.1 (DeLano Scientific LLC.). Cysteine residues are shown in red
In the homology model built using the mouse VDAC1 as template (Fig. 1), Cys2 points to the intermembrane space (IMS), whereas Cys8, which is in the same N-terminal tryptic peptide with Cys2, is located within the pore. Four of the seven cysteine residues (Cys36, Cys65, Cys122, and Cys229) are located in the loops connecting the β-strands and are therefore protruding towards IMS, as described for Cys2 (Fig. 1). Finally, Cys165 is the only cysteine exposed to the cytosol (Fig. 1).
In the modified procedure adopted for the characterization of the disulfide bonds, following the sample preparation steps (mitochondria lysis, HTP chromatography, and desalting) carried out at a pH of 6.5 to 7.0, the HTP fraction enriched in VDAC proteins was directly alkylated using IAA. The digestion step was performed using trypsin at pH of 6.5 to 7.0, conditions which are known to prevent or at least minimize disulfide scrambling [33, 35,36,37].The digestion was repeated once considering that a reduction of trypsin activity can be expected under neutral or slightly acidic conditions [33, 35,36,37]. This procedure allowed obtaining the expected tryptic peptides. Some peptide fragments originating from non-specific cleavages due to a slight increase of the chymotryptic activity of the enzyme in the adopted pH range were also detected [54]. Using the modified experimental conditions, complete sequence coverage of rVCDAC3 was obtained (Figure S1A), identifying the tryptic peptide Arg120-Lys128 and the non-specific peptides Leu114-Asp132 and Val129-Trp141, which contain the previously undetectable Lys115-Phe123 and Gly140-Trp141 stretches [23]. Similarly to what was done in the case of rVDAC2 [34], monoisotopic m/z of potential peptides containing or linked by disulfide bridges in rVDAC3 were calculated using GPMAW 9.5 software. Subsequently, pLink2 and pLabel software were used to perform a preliminary search [51] to find possible peptides linked by intramolecular disulfide bonds and to visualize the MS/MS spectra [51], respectively. The raw data were then analyzed by Xcalibur v.3.0.63 software and processed by PEAKS X-pro and MaxQuant software. Furthermore, the MaxLynx algorithm of MaxQuant [53] was used to detect and localize peptides linked by intermolecular disulfide bonds by searching for the disulfide bridge as a cross-linker. Finally, the spectra of interest were fully annotated by manually searching the full-scan mass spectrum (in high resolution) and the corresponding MS/MS spectrum (in low resolution).
Following this methodology, three intramolecular and seven intermolecular disulfide bonds between rVDAC3 with rVDAC1 and rVDAC2 isoforms were uniquely characterized, as shown in Tables 1 and 4. Furthermore, evidence was obtained for the existence of two additional intramolecular disulfide bonds between Cys2/Cys8 with Cys36 and Cys122, although these identifications were not supported by MS/MS spectra (Table 3). A map indicating the regions of the protein covered by disulfide-linked peptides is shown in Figure S1B.
The identification of the peptides in Tables 1, 3, and 4 is unequivocally supported by the high-resolution measurements, which agreed within 2 ppm with the theoretical values calculated for the disulfide-bonded peptides, and by the MS/MS spectra, in which an almost complete series of y- and b-fragments and other disulfide marker fragments for the proposed structures were observed.
It should be noted that although the three VDACs, present together in the analyzed protein mixture, share large stretches of sequence, the peptides linked by disulfide bonds are unique to each isoform as clearly visible in the Clustal Omega sequence alignment (https://www.ebi.ac.uk/jdispatcher/msa/clustalo) (Figure S2).
To confirm the results obtained and the reproducibility of the experimental data produced, the MS analysis was repeated on a second sample (biological replicate) purified from another set of rat liver mitochondria and subjected to the same procedure (data not shown).
Intramolecular disulfide bondsHRMS analysis of the sample led to the identification of a disulfide bridge in the N-terminal domain between the cysteines 2, in the acetylated form, and 8 (Table 1, peptide 1). The structure of the peptide is supported by the full-scan mass spectrum and the corresponding MS/MS spectrum of the doubly charged molecular ion at m/z 614.2549 of the N-terminal tryptic peptide (Table 1) containing the disulfide bond C2-8 reported in Fig. 2. A doubly charged molecular ion due to the same fragment with both cysteines in the carboxyamidomethylated form and Cys2 acetylated was also detected at m/z 672.2845 (Table S1, peptide 1, and Figure S3).
Table 1 Intramolecular disulfide bridged peptides in VDAC3 from Rattus norvegicusFig. 2
MS/MS spectrum of the doubly charged molecular ion at m/z 614.2549 (calculated 614.2547) of the tryptic peptide 1 (Table 1) of rVDAC3 with the cysteine residues 2 − 8 linked by disulfide bridges. In the peptide sequence, the internal fragments between the disulfide-linked cysteines are shown with yellow dashed lines (b-series) and blue dashed lines (y-series). Fragment ions originated from the neutral loss of H2O are indicated by an asterisk. Fragment ions originated from the neutral loss of NH3 are indicated by two asterisks. Disulfide-bond-specific fragment ions (cleaved at sulfur-carbon bonds) are indicated by § and in red. The inset shows the full scan mass spectrum of the molecular ion
A comparison of the absolute intensities of the doubly charged molecular ion of disulfide-bonded peptide 1 (Table 1) and that of the doubly charged molecular ion of peptide 2 (Table 2) provides a rough estimate of the relative abundance of the two cysteine forms and indicates that the disulfide-bonded fragment 1 is predominant over the reduced one, which appears to be present only in trace amounts (Table 2).
Table 2 Comparison of relative molecular ions intensities of peptides containing disulfide bridged cysteines and corresponding peptides containing carboxyamidomethylated cysteinesA disulfide bond involving Cys36 and Cys65 is demonstrated by the quadruply charged molecular ion of peptide 2 (Table 1) at m/z 877.6610, where the Tyr62-Lys74 fragment contains a missed cleavage. The full scan mass spectrum and the corresponding MS/MS spectrum are shown in Fig. 3. Furthermore, this identification is also confirmed by the detection of the triply and quadruply charged molecular ions of peptide 3 (Table 1) at m/z 1072.8252 (Figure S4A) and 804.8724 (Figure S4B), respectively, and by their MS/MS spectra.
Fig. 3
MS/MS spectrum of the quadruply charged molecular ion at m/z 877.6610 (calculated 877.6606) of tryptic peptide 3 (Table 1) of rVDAC3 with cysteine residues 36 and 65 linked by a disulfide bridge. The fragments with intact disulfide bond are reported in the MS/MS spectrum. Fragments from peptide (A) and peptide (B) are indicated in blue and green, respectively. Fragment ions originated from the neutral loss of H2O are indicated by an asterisk. Fragment ions originated from the neutral loss of NH3 are indicated by two asterisks. The inset shows the full scan mass spectrum of the molecular ion
The cross-linked and non-cross-linked peptides are present in comparable amounts as indicated by the relative intensities of the quadruply charged molecular ions of the fragments 3 and 6 (Table 2) compared to those of the doubly charged molecular ions of fragments 4, 5, and 7 (Table 2); although it should be noted that this assessment is very approximate because, due to the very different structure and mass of the two peptides, they certainly have different ionization efficiency values.
In the predicted three-dimensional conformation of rVDAC3 (Fig. 1), Cys36 and Cys65, located in the loops exposed to oxidative IMS, are in a suitable position to form a disulfide bridge. A different case occurs for Cys36 and Cys165. Indeed, based on the predicted structural model built by homology from mouse VDAC1 (Fig. 1), Cys165 is the only cysteine that faces the cytosol and therefore could not be able to bind via a disulfide linkage with Cys36 residue located in the IMS, because the two residues are extremely distant. Despite this observation, our results show that Cys36 and Cys165 are joined by a disulfide bond, as unequivocally demonstrated by the detection of the quadruply charged molecular ion at m/z 801.1125 (Table 1, peptide 4, and Fig. 4).
Fig. 4
MS/MS spectrum of the quadruply charged molecular ion at m/z 801.1125 (calculated 801.1120) of tryptic peptide 4 (Table 1) of rVDAC3 with cysteine residues 36 and 165 linked by a disulfide bridge. The fragments with intact disulfide-bond are reported in the MS/MS spectrum. Fragments from peptide (A) and peptide (B) are indicated in blue and green, respectively. Fragment ions originated from the neutral loss of H2O are indicated by an asterisk. Fragment ions originated from the neutral loss of NH3 are indicated by two asterisks. The inset shows the full scan mass spectrum of the molecular ion
Consequently, it should be assumed that the reconstructed three-dimensional structure of rVDAC3 (Fig. 1) might be incorrect or, more likely, the Cys36-Cys165 bond is an intermolecular disulfide bond formed between two different rVDAC3 molecules, whose spatial arrangement, however, cannot be derived from the present data.
The carboxyamidomethylated form of Cys165 was also identified (Table 2, peptide 10), and comparison of the absolute intensities of the corresponding molecular ions (Table 2, peptides 8–10) indicates that the disulfide bridge form (Table 2, peptides 8 and 9) is present in much lower amounts than the reduced one.
Evidence for the formation of two additional intramolecular disulfide bonds was provided by the detection of the molecular ions of the corresponding peptides, the exact masses of which were in excellent agreement with the calculated values (Table 3).
Table 3 Intramolecular disulfide bridged peptides in VDAC3 from Rattus norvegicus not supported by MS/MSEvidence for the formation of a disulfide bond between Cys2 or Cys8 and Cys122 is provided by the detection of the doubly charged molecular ion of peptide 1 (Table 3), at m/z 1063.4462. In this peptide, one of the two cysteines present in the N-terminal portion of rVDAC3 forms a disulfide bond with Cys122, while the other cysteine is carboxyamidomethylated, indicating that it is present in a reduced form in the native protein. No MS/MS was obtained corresponding to this molecular ion, probably due to its low absolute intensity, and thus, it was not possible to identify which of the two N-terminal cysteines is bound to Cys122. Likewise, the detection of a triply charged molecular ion at m/z 1073.1184 corresponding to peptide 2, Table 3, indicates the formation of a disulfide bond between Cys2 or Cys8 and Cys36. Also, for this molecular ion, no MS/MS spectrum was acquired, most likely for the same reasons seen above. The low absolute intensities of these two molecular ions could be due to the presence of a very low amount of the corresponding peptides in the analyzed mixture. However, when evaluating these results, one should take into account that if one of the two N-terminal cysteines is linked to Cys122 or Cys36, the second one could be in the reduced state in the native protein, thus being carboxyamidomethylated, as occurs in the detected peptides, but it could also be in another oxidation state, such as trioxidized to sulfonic acid or involved in another disulfide bond with another cysteine-containing peptide. While the possibility that the second cysteine is trioxidized can be excluded since the molecular ions corresponding to peptides 1 and 2 (Table 3) with a trioxidized cysteine were not present, the formation of a disulfide bond with an unknown cysteine-containing peptide cannot be excluded.
Intermolecular disulfide bondsMass spectral data confirmed the existence of supramolecular structures of VDACs due to the formation of intermolecular disulfide bonds. In this paper, we focused on the analysis of oligomeric structures between rVDAC3 and the other two isoforms (see Figures S5 and S6 for rVDAC1 and rVDAC2 predicted structures, respectively). This investigation led to the characterization of five disulfide bonds between rVDAC3 and rVDAC1, and two S–S bonds between rVDAC3 and rVDAC2 (Table 4).
In detail, the presence of the quadruply charged molecular ion at m/z 1001.2056 (Table 4, peptide 1, and Fig. 5) allowed us to demonstrate that Cys36 contained in the tryptic peptide S35CSGVEFSTSGHAYTDTGK53 of rVDAC3 is linked to Cys127 of the tryptic peptide E121HINLGCDVDFDIAGPSIR139 of rVDAC1.
Table 4 Intermolecular disulfide bridged in VDACs from Rattus norvegicusFig. 5
MS/MS spectrum of the quadruply charged molecular ion at m/z 1001.2056 (calculated 1001.2036) of tryptic peptide 1 (Table 3) with cysteine residues 127 of rVDAC1 and 36 of rVDAC3 linked by a disulfide bond. Fragments from peptide (VDAC1) and peptide (VDAC3) are indicated in blue and green, respectively. Fragment ions originated from the neutral loss of H2O are indicated by an asterisk. Fragment ions originated from the neutral loss of NH3 are indicated by two asterisks. The inset shows the full scan mass spectrum of the molecular ion
The same cysteine residue 127 of the rVDAC1 is also engaged in a disulfide bond with Cys165 of rVDAC3 as documented by the quadruply charged molecular ion at m/z 835.4053 (Table 4, peptide 2, and Figure S7).
Furthermore, Cys232 of rVDAC1, included in the tryptic peptide Y225QVDPDACFSAK236, forms disulfide bonds with cysteines 36, 65, and 165 of rVDAC3.
Indeed, the disulfide bond between Cys232 and Cys36 (peptide sequence T33KSCSGVEFSTSGHAYTDTGK53) is demonstrated by the detection of the quadruply charged molecular ion at m/z 876.6406 (Table 4, peptide 3); the quadruply charged molecular ion at m/z 730.1049 (Table 4, peptide 4) documents the formation of the disulfide bond between Cys232 and Cys65 (peptide sequence Y62KVCNYGLIFTQK74); finally, the quadruply charged molecular ion at m/z 653.5551 indicates the formation of a disulfide bridge with Cys165 (peptide sequence L164CQNNFALGYK174) (Table 4, peptide 5). The corresponding full scan and MS/MS spectra are shown in Figures S8, S9, and S10, respectively.
The two disulfide bonds between cysteines 36 (peptide sequence S35CSGVEFSTSGHAYTDTGK53) and 65 (peptide sequence V64CNYGLIFTQK74) of rVDAC3 with Cys48 in the tryptic segment S47CSGVEFSTSGSSNTDTGK65of rVDAC2 were supported by the detection of the respective quadruply charged molecular ions at m/z 946.1467 (Table 4, peptide 6, and Figure S11) and 784.1083 (Table 4, peptide 7, and Figure S12).
In order to have a rough estimate of the relative abundance of the cross-linked and non-cross-linked fragments, a comparison of their relative intensities was performed (Table 5). The results indicate that the disulfide cross-linked peptide comprising Cys127 of rVDAC1 and Cys36 of rVDAC3 (Table 5, peptide 1) is present in a very low amount. Also, cross-linked fragments between Cys48 of rVDAC2 and both cysteines 36 and 65 of rVDAC3 (Table 5, peptides 12 and 14) are detected in a very low amount. In contrast, the disulfide cross-linked peptide between Cys232 of rVDAC1 and Cys65 of rVDAC3 and the corresponding reduced peptides are detected in comparable amounts (Table 4, peptide 9). The other identified cross-linked fragments involving peptides of both rVDAC1 and rVDAC3 are found in low amounts (Table 5, peptides 4, 6, and 11).
Table 5 Comparison of relative molecular ions intensities of peptides containing disulfide-bridged cysteines and corresponding peptides containing carboxyamidomethylated cysteinesComparison of redox pattern of cysteines in rVDAC2 and rVDAC3The oxidative pattern of cysteines in rVDAC3 obtained in the present investigation was compared with that previously obtained for rVDAC2. The comparison is summarized in Fig. 6. It is very interesting to note that, in the two sequences, the homologous cysteine residues are in the same oxidation state. In particular, cysteines 2 and 8 of rVDAC3, like cysteines 9 and 14 of rVDAC2 [34], which are all located in the N-terminal α-helix region, are linked by disulfide bonds (Figs. 1 and S6). This finding suggests that at least one disulfide bridge at the N-terminus of VDAC2 and VDAC3 isoforms is required because it may have a specific function. Given its location, this disulfide bond may be indispensable to keep the N-terminal α-helix close to the channel opening in the IMS in order to regulate ions and metabolites exchange [32].
Fig. 6
Comparison of redox pattern of VDAC2 and VDAC3 from Rattus norvegicus. The N-terminal α-helix and the β-strands are shown in blue and pink, respectively. The internal loops, exposed to the intermembrane space, are indicated with I. The outer loops, exposed to the cytosol, are indicated with O
In rVDAC3, Cys2 and Cys8 were also identified in the carboxyamidomethylated form (Figure S3, Table S1, peptide 1), even if comparison of the absolute molecular ion intensities of the oxidized disulfide-bridged versus the reduced one indicates that the latter is present in a very small amount (Table 2).
Similarly, cysteines 36, 65, and 165 of rVDAC3 as well as the homologous cysteines 48, 77, and 104 of rVDAC2, which are located in the loops connecting the β-strands (Figs. 1 and S6), are oxidized to both sulfonic acid (Table S4, peptides 1–6, and Figures S13-S15) and disulfide bridges (Table 1, Figs. 3, 4, and S4). Furthermore, the cysteines 36, 65, and 165 (Table S1, peptides 2, 3, 4, 6, and 7, and Figures S16-S18) are also detected in the reduced form in considerable amounts (Tables 2 and S5).
Cys122 of rVDAC3, located in a loop region exposed to IMS (Fig. 1), is homologous to Cys134 of rVDAC2, which is linked to Cys139 by a disulfide bond particularly resistant to reduction [34]. A cysteine residue homologous to Cys139 of rVDAC2 does not exist in rVDAC3. As reported in Fig. 6, in this study Cys122 is detected as carboxyamidomethylated (Table S1, peptide 5, Figure S19), indicating that it is present partially in the reduced form in the native protein, but also with the variable “Half of a disulfide bridge” modification (Table S6, Figure S20), suggesting that it is engaged in a disulfide bond, although the cysteine residue to which it is bound is unknown. The predictable peptides corresponding to the formation of a disulfide bond with another rVDAC3 molecule or another rVDAC isoform were not found; therefore, the disulfide bond in which Cys122 is involved cannot be defined [55].
Finally, Cys229 of rVDAC3 and its homologous Cys228 of rVDAC2, which, based on the predicted three-dimensional structure, are located in the IMS-exposed region (Figs. 1 and S6), are always predominantly oxidized to sulfonic acid [23] (Table S4, peptide 7, and Figure S21), while the corresponding reduced forms are present only in trace amounts (Table S1, peptide 8, Table S5, and Figure S22).
The formation of intermolecular disulfide bonds helps to explain the previously observed VDAC oligomerization [56]. Oligomerization of more VDAC monomers has been proposed to allow the formation of a supramolecular larger pore, which can be used as a conduit large enough to allow the exit of DNA mt molecules or fragments [57], and to contribute to the permeability transition [58]. Multimeric pores of sufficiently large diameters could release cytochrome c from the mitochondrial IMS to the cytosol, thus triggering mitochondria-mediated apoptosis [56]. Forces and bonds involved in the VDAC oligomerization expect precise identification. Intermolecular disulfide bridges could contribute to the formation or stabilization of oligomers. A suggestion in this sense has been recently proposed in [6].
Comments (0)