
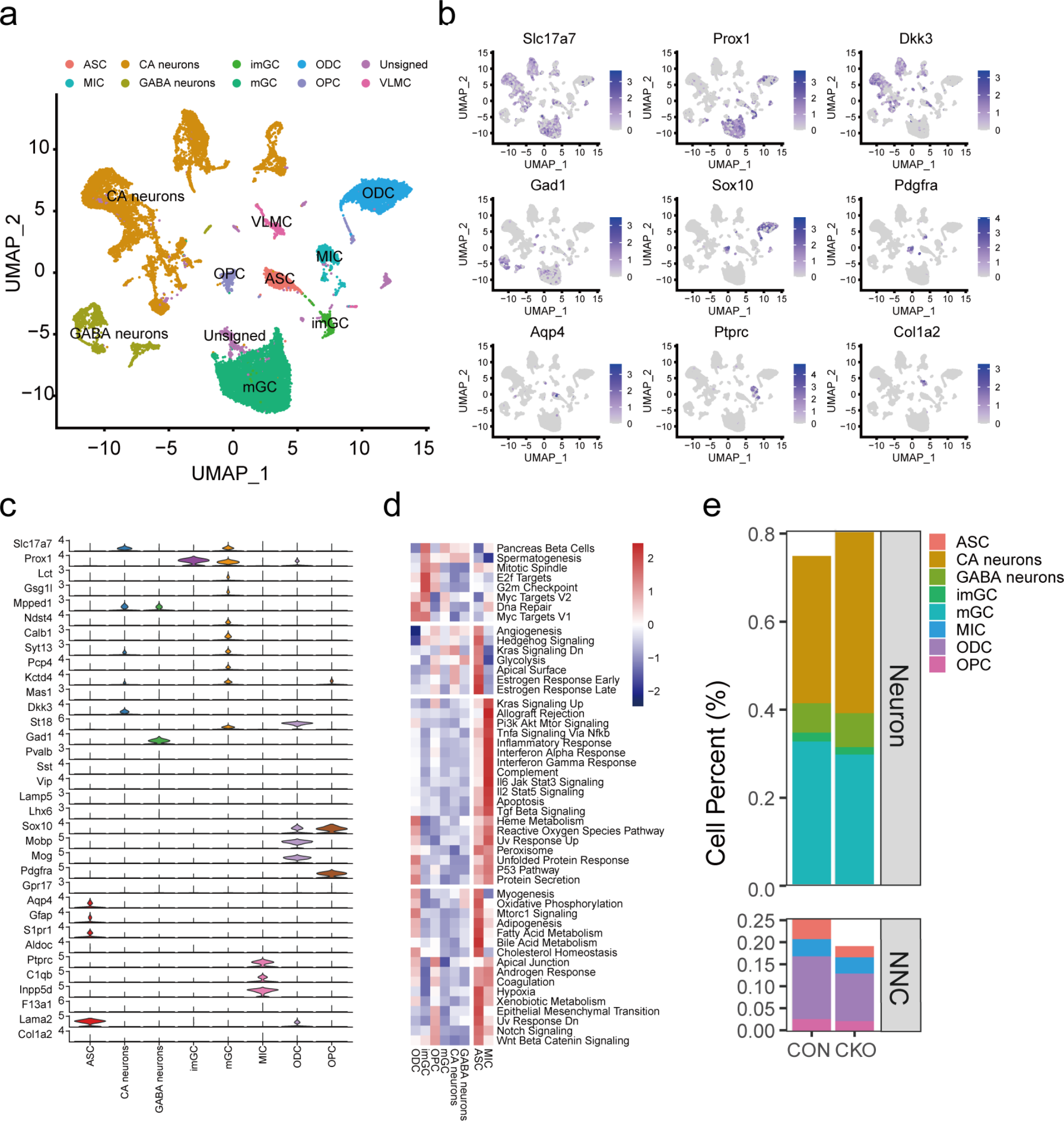
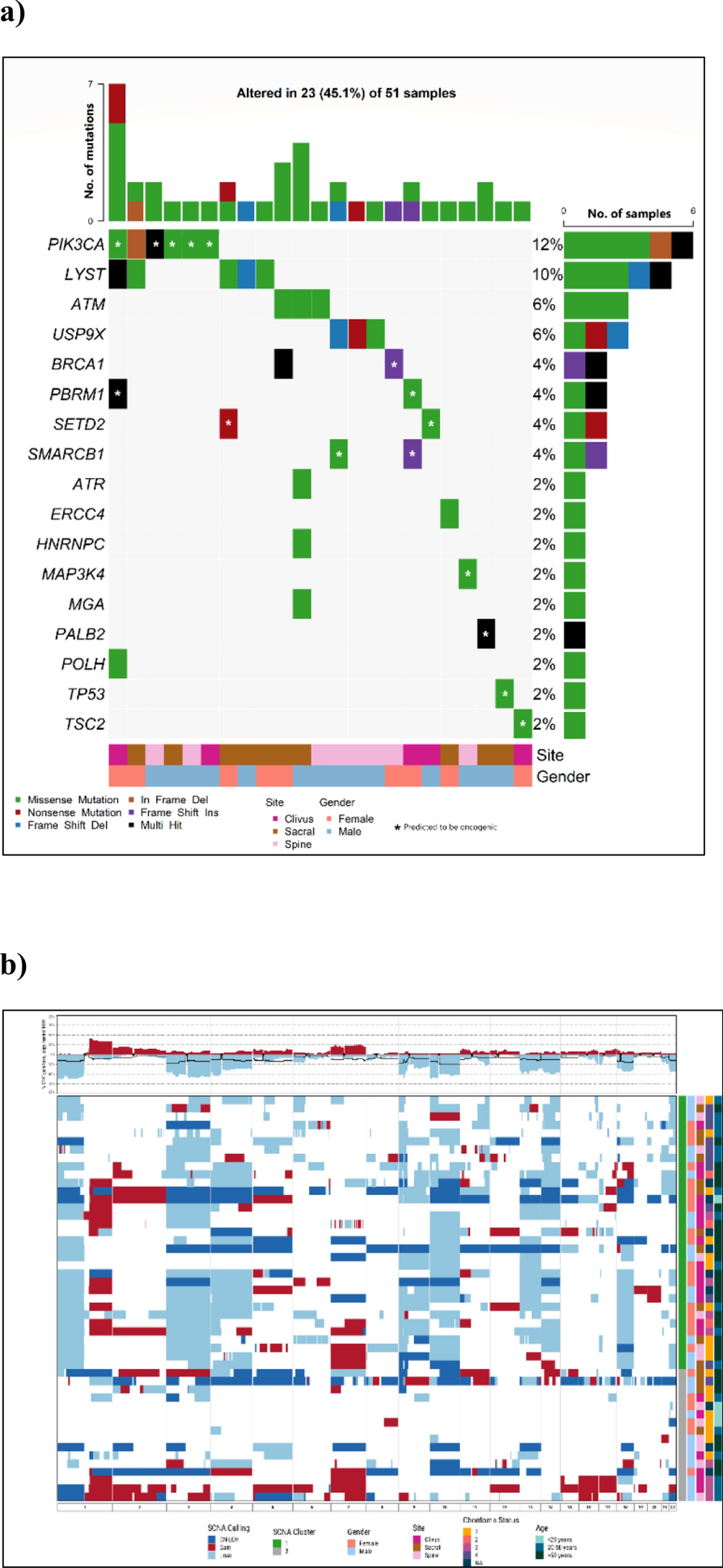
Remember me




The patient was born prematurely, at 32 weeks of gestational age, from healthy, non-consanguineous parents. His birth weight was 1790 g, the Apgar score was 10/10. An intracranial hemorrhage grade III was diagnosed. Clinical reports showed that the patient presented with motor delays, developmental delays, autism spectrum disorder, hypotonia, and small genitalia. His parents also reported visual impairments, feeding and eating problems, as well as sleep disorders. Phenotypically, the patient presented with a prominent forehead and eyelashes, downward slanting eyes, malformed ears, wide nasal bridge, broad and long philtrum, large mouth with thick lower vermillion, pointed chin and widely spaced teeth (Fig. 1A, B), all well-defined characteristics described in a cohort of 78 Helsmoortel–Van der Aa patients [2, 4] (Additional file 1: Table S6). At the age of 2.5 years, he developed an upper respiratory tract infection complicated with hepatitis and seizures. He was transferred to ICU where supportive treatment and plasmapheresis were started. Liver biopsy showed extensive necrosis of parenchyma and moderate cholestasis. MRI showed diffuse cortical atrophy of the brain parenchyma, marked reduction in volume of white matter as well as gliosis in both frontal and temporoparietal lobes that could indicate the sequelae of acute hepatic encephalopathy. He developed refractory generalized epilepsy and received a combination treatment of antiepileptic drugs, e.g., carbamazepine, oxcarbamazepine, levetiracetam, clonazepam, clobazam and topiramate. During his lifespan, he underwent two liver transplantations and received immunosuppressants. Following the second liver transplant, at the age of six years and three months old, the child passed away because of multiple organ failure. An autopsy was performed, and various tissue samples were donated with informed consent. Molecular testing had indicated that the patient was negative for any inheritable metabolic disorders. Whole-exome sequencing (WES) of the patient’s blood revealed a heterozygous de novo duplication of adenine at position 1676 in the ADNP gene at position chr20:50,893,037-50,893,039 (RefSeq isoform ENST00000621696.5 Human GRCh38/hg38). The mutation was confirmed by Sanger sequencing (Fig. 1C). It converts the histidine (His) residue at position 559 to glutamic acid (Gln), leading to a frameshift mutation with a premature stop codon two amino acids downstream (Fig. 1D, E).
Fig. 1
Identification of a heterozygous de novo mutation in the ADNP gene. (A) Facial photograph of the six-year-old child (https://www.adnpfoundation.org/). (B) Schematic representation of the clinical manifestation of the patient with Helsmoortel–Van der Aa syndrome, including autism, severe ID, and epilepsy. (C) DNA sequencing chromatogram of control and patient alleles, confirming a heterozygous nucleotide duplication (c.1676duplA) in the ADNP gene, (D) replacing the histidine at residue 559 with glutamic acid with a frameshift of two amino acids and introduction of a stop codon (p.His559Glnfs*3). (E) ClustalW alignment across multiple species of ADNP amino acids 520–580. Almost all residues of the ADNP protein are highly conserved amongst vertebrates. The arrow (↓) indicates the species-conserved histidine (H) residue, which is altered in the patient to a glutamic acid (Q) residue. The asterisk (*) indicates positions which have a single, fully conserved residue. A colon (:) indicates conservation between amino acid residues of similar properties
Cerebellar tissue, known for its highest ADNP expression [35], allowed to validate the presence of ADNP mRNA and protein in autopsy material by performing an expression analysis using real time reverse-transcription PCR (RT-PCR) and Western blotting. To investigate wild-type ADNP mRNA levels, we designed a primer set at the 3’ region of exon 6 (corresponding to the C-terminal portion of the protein). Here, a significant two-fold increase in the total ADNP levels was observed in the patient compared to the control subject (p = 0.0001; ***), consistent with findings in our RNA sequencing described below (Fig. 2A). Attempts to quantify the 5’ end of the transcript were not successful, suggesting partial mRNA degradation. At the protein level, we tested endogenous ADNP levels in the human brain using extensively validated C-terminal and N-terminal ADNP antibodies [29]. We were able to detect wild-type ADNP levels (150 kDa) in the control brain, but not in the patient using both antibodies (Fig. 2B, C). To investigate the co-expression of the full length and mutant protein, we co-transfected wild-type and p.His559Glnfs*3 mutant N-DYKDDDDK (Flag®) expression vectors in HEK293T cells. Co-expression of wild-type and mutant ADNP demonstrated the presence of the wild-type protein (150 kDa) together with a truncated mutant protein (63 kDa) using an N-terminal antibody, mimicking the expected expression in the patient. C-terminal antibody incubation resulted in the detection of the wild-type ADNP (150 kDa) exclusively. Together, these findings confirm a molecular weight of ADNP (150 kDa), above its calculated molecular weight of 123 kDa, but show instability of the protein in post-mortem brain material of the patient.
Fig. 2
ADNP expression analysis of the cerebellum. (A) RT-PCR showing a significant increase in ADNP mRNA levels in the patient cerebellum compared to an age-and sex matched control subject (***p = 0.0001; unpaired student T-test). Gene expression values were normalized with three stable reference genes, i.e., β-Actin (BACT), β-2-Microglobulin (B2M), and Ubiquitin C (UBC). (B) ADNP protein expression analysis using an N-terminal antibody. Western blotting showed the presence of wild-type ADNP (150 kDa) in overexpression lysates with presence of the truncated protein. However, expression was absent in the patient as compared to the control, where wild-type ADNP could be visualized. (C) ADNP protein expression analysis using a C-terminal antibody. Western blotting showed the presence of wild-type ADNP (150 kDa) in overexpression lysates, as well as in the control subject, but not in the patient. GAPDH was used as a loading control for normalization
To study the molecular impact of the patient mutation, we performed in silico modeling of the wild-type ADNP protein (UniProt; Q9H2P0) and p.His559Glnfs*3 mutant using AlfaFold. Here, the structure of the wild-type protein demonstrated the DNA-binding homeobox domain in proximity to the bipartite NLS sequence, whereas the neuroprotective NAP motif resides at the surface of the protein, being partially occluded by flexible intrinsically disordered regions (IDRs) and low-complexity regions (LCRs) located near the C-terminus, suggestive for a role for protein–protein interactions [36, 37]. Moreover, the eIF-4E binding motifs and the glutaredoxin active site are centrally positioned in the core of the wild-type protein, assembling several of its zinc finger motifs (Fig. 3A). The p.His559Glnfs*3 mutant truncates the NLS region, impairing nuclear transport [11]. Moreover, downstream protein domains, including the DNA-binding homeodomain and the HP1 binding motif are also lost as a result of the truncating mutation. Overall, the p.His559Gln*3 mutant lacks some of the IDRs but has a similar structural confirmation compared to the wild-type protein (Fig. 3B). Subsequently, we examined stable ADNP protein levels in several subcellular compartments including the cytoplasm, nucleus with chromatin-enriched proteins, and the cytoskeleton in HEK293T overexpression lysates. In the cytoplasm, we detected wild-type (150 kDa) and mutant (63 kDa) ADNP using an N-terminal antibody showing no significant difference in expression levels (p = 0.71; ns). In the chromatin-bound fraction, we visualized the wild-type and mutant protein with a significant decrease of mutant protein levels (p = 0.03; *). Moreover, we demonstrated the expression of mutant and wild-type ADNP in the cytoskeletal protein fraction. However, we did not observe a significant difference (p = 0.42; ns) in the expression of the mutant compared to the wild-type protein (Fig. 3C).
Fig. 3
The ADNP patient mutation impairs expression in the chromatin-enriched protein fraction. (A) 3D protein structure representation of the wild-type ADNP glutaredoxin active site (pink), NAP octapeptide sequence (fuchsia), eIF-4E interaction motif (blue), nuclear localization signal (dark cyan), homeobox domain (blue violet), and HP1 interaction motif (purple). The NAP domain (fuchsia) presents at the surface of the protein. (B) The nuclear localization signal-truncating p.His559Glnfs*3 mutant shows loss of the HP1-binding motif and DNA homeobox domain. (C) N-terminal ADNP detection in different subcellular compartments normalized to their protein fraction-specific loading controls. Detection of wild-type N-DYKDDDDK (Flag®)-tagged ADNP shows a molecular weight of 150 kDa. The p.His559Glnfs*3 mutant showed a lower molecular weight of 63 kDa. Cytoplasmic enrichment shows expression of wild-type ADNP (150 kDa) and the mislocalized p.His559Glnfs*3 mutant (63 kDa) with no difference in expression (p = 0.71; ns). Chromatin-enriched fraction demonstrated partial loss of mutant ADNP levels compared to wild-type ADNP, showing a dramatic decrease in expression (p = 0.03; *). Cytoskeletal fraction is enriched for wild-type ADNP and the p.His559Glnfs*3 mutant, with no significant difference in expression (p = 0.42; ns). GAPDH (cytoplasmic fraction), histone H3 (chromatin-bound fraction), and β-actin (cytoskeletal fraction) were used as loading controls. Statistical analysis of the subcellular fractionation immunoblots was performed using an unpaired two-tailed student T-test, assuming equal variances
Genome-wide methylation analysis of the cerebellum demonstrates abnormalities of the cytoskeleton and autophagy together with an aberrant transcription factor function of ADNP during developmentAs methylation signatures are robust and even conserved in ancient DNA [38], we decided to start our exploration by performing an EPIC BeadChip array on the cerebellum of the died ADNP patient and an age-matched control brain. Here, we show enrichment of 6289 CpG probes with a minimum 20% difference in methylation in the ADNP patient compared to the control. Specifically, we identified 2394 CpG probes showing hypermethylation (Δβ > 0.2), whereas a vast amount of 3895 CpG probes were hypomethylated (Δβ < −0.2). In addition, 1547 hypermethylated gene probes could be annotated to 1162 genes, while 2500 hypomethylated gene probes were associated with 1842 genes (Additional file 3: Data S1), indicating a Class I episignature [18, 19], extending findings from peripheral blood to the human brain for the first time (Fig. 4A). Next, we confirmed a selection on genes prioritized for methylation in the 5’UTR, 3’ UTR and transcription start site (TSS) together with associations to autism or other Helsmoortel–Van der Aa syndrome-related clinical features. We selected the hypermethylated genes OTX2, SLC25A21, and DNAJ6 and the hypomethylated genes COL4A2, MAGI2, and CTNND2 for pyrosequencing. Here, we could confirm a higher percentage of CpG methylation in the patient for OTX2 (56%), SLC25A21 (86%), and DNAJ6 (85%) compared to the control subject. Respectively, we could also demonstrate a lower percentage of CpG methylation in the patient for COL4A2 (1%), MAGI2 (2%), and CTNND2 (3%) (Fig. 4B). Next, we performed functional annotation of the hyper- and hypomethylated genes using Metascape. Enriched biological processes and GO terms included actin filament-based processes, cell adhesion, nervous system development, muscle contraction, brain development, the WNT signaling pathway, regulation of membrane potential, and synaptic transmission amongst others (Fig. 4C). Functional enrichment analysis for protein–protein interactions was predicted for ADNP using the STRING database. We identified four suggested interactions of ADNP with WDFY3, UBR5, FAT1, and NFIA, which play a role in autophagy of the mitochondria, protein ubiquitination, macro-autophagy, autophagosome and autolysosome formation (Fig. 4D). Given the role of Adnp as a putative transcription factor [35, 39], we performed a transcription factor enrichment of both hyper- and hypomethylated genes. Here, we identified a module of 44 co-expressed genes, which were subsequently inserted in CytoScape using the IRegulon function for TF enrichment (Additional file 4: Data S2). We observe a stronger enrichment of TFs associated with hypomethylated genes (red) than hypermethylated genes (blue) and shared TFs (green). Among the upregulated TFs associated with hypomethylated genes presented pluripotency and cell fate-determining genes such as POU2F1, TEAD2, SOX1/4, GATA1/2/3/5/6, PAX4/6, NANOG, and NEUROD1, as well as chromatin modifiers like YY1, SIN3A and ADNP itself. On the other hand, the downregulated TF cluster associated with hypermethylated genes was also enriched for PAX and SOX-related genes, indicating abnormal lineage specification of neuronal progenitor cells. The shared TF cluster showed presence of HNF1A, a gene controlling expression of several liver-specific genes (Fig. 4E). Our genome-wide cerebellar methylation analysis indicates strong molecular evidence for a deregulated function of ADNP as a transcription factor, impacting lineage specification and genes implicated in brain development.
Fig. 4

ADNP methylation signature in the juvenile post-mortem cerebellum. (A) Genomic scatter plot indicating the hypermethylated genes (Δβ > 0.2) of the patient (red), the hypomethylated genes (Δβ < −0.2) of the patient (blue). The chromosomal positions of the genes are shown on the x-axis. (B) Pyrosequencing confirmation of a subset of hyper- and hypomethylated genes. Hypermethylated genes, e.g., OTX2, SLC25A21 and DNAJ6, show increased CpG methylation in the patient, whereas hypomethylated genes, e.g., COL4A2, MAGI2 and CTNND2, present with a nearly absent percentage of CpG methylation. (C) Metascape functional annotation of biological processes. Hyper- and hypomethylated genes cluster in associated processes such as the actin cytoskeleton and nervous system developmental disorder amongst others. (D) Predictive String v11.5 protein–protein interaction analysis of ADNP. The proteins are indicated as nodes with interconnecting lines representing the interaction. ADNP is surrounded by protein regulating specific autophagy-related processes and protein ubiquitination. (E) Transcription factors (TFs) enriched in patient cerebellum of hyper- and hypomethylated gene co-expression. TFs associated with hypermethylated genes are represented in blue, while the TFs associated with the hypomethylated genes are depicted in red. TFs shared amongst the overlapping genes are shown in green. ADNP was identified as the top transcription factor controlling the hypomethylated genes (black box)
RNA sequencing substantiates downregulation of the WNT signaling pathway and autophagy defects in cerebellar autopsy tissueTo determine differential expression beyond methylation differences, we performed bulk transcriptome sequencing of cerebellar tissue of the ADNP autopsy. As RNA is much less stable over time, we first performed an extensive quality control by evaluating total RNA purity and integrity (see experimental methods). Using bulk mRNA sequencing, we determined the gene ratio (patient/control) using the NOISeq algorithm, a non-parametric method for comparing samples without biological replicates, reporting the log2-ratio of the two conditions (M) and the value of the difference between the conditions (D) [40]. We tested for differential expression across all 7659 genes that appeared in our data set (Additional file 5: Data S3). In line with the observation of an excess of hypomethylated CpG probes, we observed an excess of upregulated genes. Using a significance cut-off equivalent to, p value < 0.05, FDR = < 0.05, and a biologically meaningful (M-value) log2FC > 0.5, we found 514 downregulated and 1520 upregulated genes with differential expression (Fig. 5A). Gene expression alterations in the ADNP cerebellum were notable with the majority of genes presenting with an M-value < 5. Gene ontology (GO) enrichment revealed downregulation of glutamatergic synaptic transmission, abnormal cardiac muscle cell conductivity, and nervous system development, whereas cytoskeleton dynamics were upregulated. A remarked enrichment of immune system-related responses was observed that are potentially related to the patient’s immunosuppressant treatment (Fig. 5B). We confirmed a selected set of genes with RT-PCR, including the RNA-methylation gene METTL3 (p = 0.005; **), autophagy inducer BECN1 (p < 0.0001; ****), and WNT signaling ligand CTNNB1 (p = 0.001; **) (Fig. 5C). To better interpret the differential expression in the ADNP brain, we compared the transcriptome analysis of the autopsy with the differential expression observed in immortalized LCLs of multiple patients with different ADNP mutations. We tested for differential expression across approximately 10,000 protein-coding transcripts that appeared in our data set (Additional file 6: Data S4). Using the exact cut-off criteria as in the autopsy, we found 1730 downregulated and 3278 upregulated genes with differential expression, indicating that the ADNP mutations rather induce gene upregulation (Fig. 5D). Fast Gene Set Enrichment Analysis (fgsea) identified similar molecular pathways as identified in the autopsy (Fig. 5E). We confirmed a subset of five genes with RT-PCR, including the heterochromatin marker and ADNP-interacting gene CBX3 (p = 0.01; *), WNT signaling member WNT10A (p = 0.003; **), actin-cadherin mediator CTNNAL1 (p = 0.003; **) as well as nonsense mediated decay members SMG5 (p = 0.0002; ***) and UPF3B (p = 0.005; **) (Fig. 5F).
Fig. 5
Cerebellar and lymphoid gene expression changes are associated with different ADNP mutations. (A) Volcano plot of differentially expressed genes (DEGs) in the ADNP cerebellum using the NOISeq algorithm, representing the effect size M (log2 ratio) and D (difference between conditions) values. The DEG are shown in blue. (B) Gene set enrichment analysis of all DEGs in gene ontology (GO), biological processes (BP) and molecular function (MF) reveals specific Helsmoortel–Van der Aa syndrome-related pathways in the ADNP brain. (C) RT-PCR showing a significant reduction of METTL3, BECN1 and CTNNB1 mRNA levels in the ADNP cerebellum compared to the age-matched control subject. Expression values were normalized using the housekeeping genes ACTB, B2M and UBC. Data was analyzed using an unpaired student T-test. (D) Volcano plot of DEGs in the patient LCLs using the DESeq2 package, displaying the significance (-log10q) and effect size (log2FC). The DEG are shown in blue. (E) Functional gene set enrichment of GO and BP using differentially expressed genes in the ADNP LCLs as compared to age- and sex-matched controls. UMAP clustering of gene sets colored by standard deviation, variance, or mean fold-change in patient LCLs shows clear downregulation of the WNT, Hedgehog and Notch signaling pathways (marked in a red box), impairing proper neuronal development. Downregulated genes, blue; upregulated genes, red. (F) RT-PCR showing a significant increase of CBX3, CTNNAL1, SMG5 and UPF3B together with a significant decrease in WTN10A mRNA levels in patients versus control LCLs. Expression values were normalized using the housekeeping genes GAPDH, RPL13A and SDHA. Data was subsequently analyzed with a Mann–Whitney U test for unpaired measures
To investigate the potential impact of the ADNP mutation in the human brain, we intersected the DEGs identified in both data sets (Additional file 7: Data S5), which revealed an overlap of 241 genes between the ADNP autopsy brain and LCLs (Fig. 6A). We observed a striking resemblance for biological relevance of genes involved in endoderm specification IGFBP2 (brain, *p = 0.03; LCL, *p = 0.04), canonical WNT signaling WNT2 (brain, *p = 0.01; LCL, **p = 0.01), mitochondrial transporter SLC25A25 (brain, *p = 0.02; LCL, *p = 0.03), autophagy regulation RUBCN (brain, ****p < 0.0001; LCL, *p = 0.003), hematopoietic stem cell differentiation RUNX1 (brain, **p = 0.001; LCL, ***p = 0.001), N6-adenosine-methylation METTL3 (brain, **p = 0.005; LCL, ****p < 0.0001), and bone and teeth development BMP6 (brain, **p = 0.002; LCL, *p = 0.04) (Fig. 6B, C). In conclusion, these robust gene expression changes related to the nervous system and morphogenesis underline a regulating role of ADNP in the human brain and blood of patients, confirmed by salient pathways including the WNT signaling, autophagy, and bone development together with involvement in processes such as hematopoietic stem cell differentiation and unexpected RNA methylation.
Fig. 6
Meta-analysis of the transcriptomic signature identified in the ADNP brain and lymphoblasts. (A) Venn diagram representing the exact amount of DEGs in the human ADNP cerebellum and lymphoblastoid data sets, converging to an amount of 241 overlapping genes. (B) RT-PCR showing a significant decrease of IGFBP2, WNT2, SLC25A25 together with a significant increase in RUBCN and RUNX1 mRNA levels in patient brain and patient LCLs as compared to their age-matched controls. Note a difference in mRNA expression of BMP6 and METTL3 amongst brain tissue and LCLs. Expression values were normalized using the tissue-specific housekeeping genes (mentioned above). Data was subsequently analyzed with an unpaired student T-test (brain) or Mann–Whitney U test for unpaired measures (LCLs). (C) Correlation of DEGs from the NOIseq and DESeq2 analysis (RNA-seq) and RT-PCR confirmations with their functional cellular role. Selected genes are represented with their Log2FC and compared for overlap with RT-PCR. Arrow upwards, upregulation; arrow downwards, downregulation
Shotgun proteomics links chromatin remodeling to autophagy in the ADNP autopsy brainAs post-transcriptional regulation can further increase variation in gene expression levels [41], proteome analysis was performed by label-free quantitation (LFQ) mass spectrometry on the cerebellum to study the effect of the c.1676duplA/p.His559Glnfs*3 ADNP mutation at protein expression level. Chromatographic conditions between different runs were highly reproducible, resulting in a strong correlation between LFQ intensities and technical replicates (Additional file 8: Data S6). Overall, we detected approximately 1522 protein groups per sample under a 1% false-discovery rate (FDR) with fixed modifications of carbamidomethylation (C), deamidation (QN) and oxidation (M). Moreover, we identified 4552 proteins with more than two unique peptides, respectively 988 proteins with at least two unique peptides, and 1477 with one unique peptide. Next, we used MetaboAnalyst 5.0 to quantify differences detected in patient versus control cerebellum. Among the 2455 quality-filtered proteins, we detected 492 proteins with a differential expression (Additional file 9: Data S7), of which 224 proteins were significantly downregulated, while 268 proteins showed a significant upregulation in the post-mortem patient cerebellum (two-tailed student T-test; padj < 0.05). Next, we plotted the top 10 downregulated (represented in blue) and upregulated (represented in red) proteins identified in patient versus control brain, showing a clear upregulation of the major ADNP-interacting protein heterochromatin Protein 1 homolog beta (CBX1/HP1β), amongst others, indicating that ADNP is able to somehow affect the expression of its direct interaction partner (Fig. 7A). Subsequently, we applied immunoblot experiments to confirm the downregulation of β-catenin and BECN1 protein levels in the patient brain, in line with its decreased transcription levels. Surprisingly, we also observed differential expression of an additional autophagy marker, MAP1LC3A, in the ADNP brain consistent with aberrant autophagy defects in our transcriptome data (Fig. 7B). Clustering of the differentially expressed proteins (DEPs) in canonical pathways using IPA indicated a decreased activity of mitochondrial oxidative phosphorylation, sirtuin signaling and RhoA signaling. In contrast, IPA predicted an increase in EIF2 signaling, spliceosomal cycle and protein kinase A signaling in the patient. We also observed an enrichment of pathways with no difference in activity, including granzyme A signaling and mTOR signaling, T-helper signaling, and apoptosis (Fig. 7C). Next, we mapped all DEPs in a functional enrichment analysis and predicted possible protein–protein interactions of ADNP with the identified DEPs as well as with other biologically correlated proteins. Of particular interest, the histone deacetylase sirtuin 1 (SIRT1) in the center of the protein network was found to link various chromatin modifier proteins such as MECP2, ADNP, SMARCC2, HDAC2 including YY1, and chromatin-associating proteins such as CBX1/3, histones H1F0 and H1.2 to autophagy regulators like MAP1LC3A and LAMP1 (Fig. 7D). In this section, we showed that the proteomic landscape of ADNP brain autopsy material corroborates our transcriptome findings, e.g., upregulation of ADNP-interactor CBX1/HP1β together with a downregulation of β-catenin and BECN1, supported by abnormalities attributed to the WNT signaling pathway and autophagy.
Fig. 7
Discovery of SIRT1 interactions in the autistic brain, linking chromatin remodelers to autophagy. (A) Volcano plot of differentially expressed proteins (DEPs) in the ADNP patient cerebellum, represented by the significance (−log10(p)) and effect size (log2FC). Proteins with a significant downregulation are shown in blue and those with an upregulation in red. Note the marked difference in expression of the ADNP-interacting protein CBX1/HP1β. (B) Immunoblot confirmations of DEPs exposed to their specific antibodies. (C) Top 15-ranked canonical IPA pathways of the ADNP brain, represented as z-scored expression values and significance (−log p-value). Activated pathways are presented in orange, respectively lowered activity in blue; no activity pattern available (grey); pathways with no difference in activity (white). (D) A predictive protein–protein interaction network was generated by String Version 11.5, integrating associations of a co-expression hub identified amongst the DEPs. Proteins are represented by colored network nodes in relation to each other with SIRT1 fulfilling a centralizing role. The edges illustrate functional associations and the lines between the nodes represent the existence of evidence for associations
ADNP and SIRT1 co-immunoprecipitate with the microtubule end-binding proteins 1 and 3 (EB1/EB3)Recently, various studies identified an association between mitochondrial dysfunction, autophagy regulation, and autism spectrum disorders [42,43,44]. Similarly, our proteomic protein–protein interaction study mapped SIRT1 at the crossroads of chromatin remodelers and autophagic regulators in the ADNP autopsy brain. Besides, SIRT has been discovered to maintain genomic stability [45], to enhance synaptic plasticity [46], to suppress inflammation [47], to fulfill a neuroprotective function [48], and to positively regulate autophagy and mitochondrial function [49]. In addition, SIRT1 is known to modulate chromatin structure by activating BRG1, which is a chromatin remodeling interaction partner of ADNP in the SWI/SNF complex [9, 50]. Hence, we reasoned that ADNP and SIRT1 may share common regulatory partners in chromatin remodeling and microtube dynamics that regulate autophagy. To further validate a direct protein interaction of ADNP and SIRT1 in the human brain, co-immunoprecipitation (co-IP) experiments were performed. However, due to the instability of the ADNP protein, co-IP experiments were not successful. Therefore, we alternatively demonstrated the subcellular localization of Adnp and Sirt1 by immunostaining the cerebellum dissected from male C57BL/6JCr wild-type mice as a model for the human condition. Here, cerebellar cryosections were immunostained with primary monoclonal ADNP and SIRT1 antibodies (Cy3 red fluorescent signal), and nuclei were counterstained with DAPI (blue). Adnp expression was predominantly detected in the nucleus, with occasional weak cytoplasmic signals, visualized by the overlap of the red Adnp signal and blue DAPI counterstaining. In contrast, Sirt1 was predominantly situated in the cytoplasm of Purkinje cells in the cerebellum, with occasional nuclear immunoreactivity (Fig. 8A). Indeed, an indirect interaction of ADNP and SIRT1 was shown in SH-SY5Y cells which could not be validated in human induced pluripotent stem cells (hIPSC)-derived neuronal differentiated cells [16]. Therefore, we next investigated the potential indirect interaction of Adnp and Sirt1 through the EB1/EB3 proteins in murine cerebellar brain lysates with a co-immunoprecipitation assay. During this process, we performed stringent washing steps using high detergent buffers to prevent false-positive binding. In addition, we also controlled each western blot with GAPDH, whose intensity was absent after co-immunoprecipitation of the bait protein. We observed specific co-immunoprecipitation of Adnp (150 kDa) and Sirt1 (100 kDa) in the presence of both EB1 (30 kDa) and EB3 (32 kDa) antibodies. IgG non-reactive beads were used as a negative control, showing no immunoreactivity of Adnp, Sirt1 together with EB1 and EB3 in the eluted fraction (Fig. 8B). To better understand the physical connections between Adnp (UniProt; Q9Z103) and Sirt1 (UniProt; Q53Z05), we applied a eukaryotic linear motif (ELM) analysis to unravel shared motifs (Fig. 8C). Interestingly, as partially shown before, we identified a series of common interaction motifs, including (1) SxIP motif for Adnp (aa 354–360, NAPVSIP, p = 0.01) and similar SSIP for Sirt1 (aa 440–448, VALIPSSIP, p = 0.0002), (2) SH3-domains for Adnp (i.e., aa189–195, FQHVAAP, p = 0.01) and Sirt1 (i.e., aa 506–511, PPRPQK, p = 0.001), and (3) 14–3–3 motifs for Adnp (i.e., aa 16–20, RKTVK, p = 0.004) and Sirt1 (i.e., aa 333–342, RNYTQNIDTL, p = 0.004). The presence of a SxIP motif is a unique feature ascribed to both ADNP and SIRT1, as only 42 protein have been identified by mass-spectrometry based methods to contain this conserved motif [51, 52]. Original studies have identified the microtubule-end binding proteins EB1 and EB3 as interaction partners of ADNP through its SxIP motif [16, 23, 27]. Along the same line, we predicted a physical interaction between Adnp, Sirt1 and EB1/EB3 in silico via 3D-molecular docking. Upon ranking the models according to the amount of interacting amino acids, the top 10 docking interactions were derived from ClusPro and processed in ChimeraX. Molecular docking revealed possible Adnp (blue) binding to both microtubule-end binding proteins EB1 (left, violet) and EB3 (right, pink) via amino acids 358–360, corresponding to its SxIP motif. In addition, Sirt1 was predicted to interact with EB1 (left, violet) and EB3 (right, pink) proteins through its similar SSIP motif at amino acid position 446–448 (Fig. 8D). In conclusion, our findings suggest that Adnp and Sirt1 might indirectly co-immunoprecipitate in the presence of the EB1/EB3 proteins via the SxIP motif for ADNP, respectively SSIP for SIRT1.
Comments (0)