
Remember me
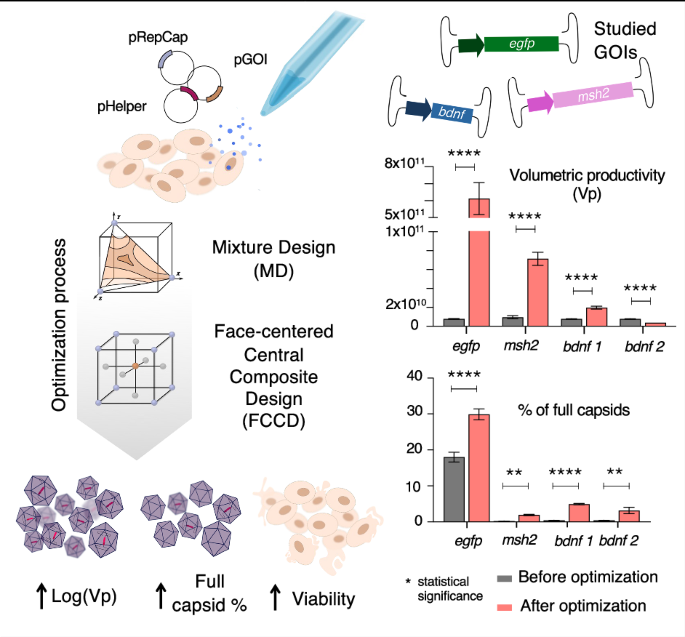
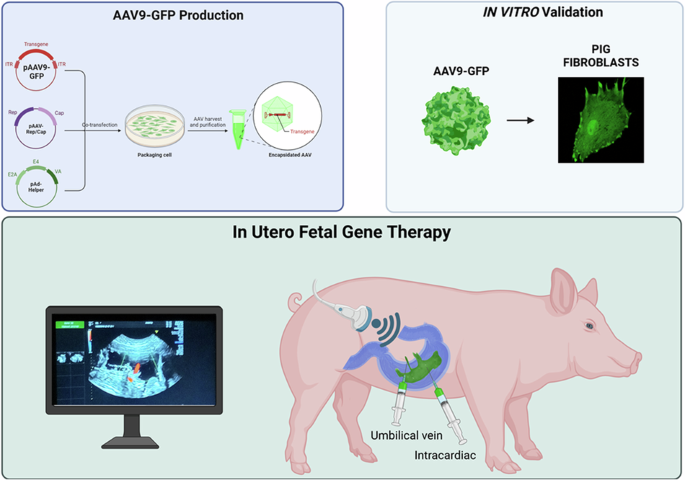
We developed a minimally invasive, ultrasound (US)-guided technique for delivering AAVs to fetal piglets in utero. This approach, akin to standard procedures in fetal medicine such as blood transfusion, can be readily adapted for clinical application (see Experimental section for details).
We performed four rounds of in-utero injections in four pregnant sows ranging from gestational age (GA) 80–108 (pig gestation is ~115 days), corresponding to the beginning of the third trimester to the end of the third trimester in humans [23].
Under direct ultrasound visualization, the operator inserted a 20-gauge spinal needle at a 45° angle relative to the ultrasound beam. The needle was guided through the maternal skin, the uterus, and into the amniotic cavity (Fig. 1a). The needle was then inserted in the umbilical vein near the placental cord insertion (Fig. 1b and Supplementary Video 1) or in the fetal heart chamber of the left ventricle (Fig. 1c and Supplementary Video 2) to deliver the virus. The entire procedure, including intubation and surgical preparation of the sow, fetal identification, and virus injection, had an average duration of 28 min. Fetuses received injections of scAAV9-CAG-GFP (n = 7) or saline (n = 2).
Fig. 1: Ultrasound-guided, in-utero fetal injection procedure in pigs.
a Representative image of the transabdominal injection. b Ultrasound image of the scAAV9-CAG-GFP injection in the fetal umbilical vein. c Ultrasound image of intracardiac scAAV9-CAG-GFP injection.
The viral doses injected into the fetuses ranged from 1.2 × 10¹² viral genomes (vg)/kg to 5 × 10¹³ vg/kg, resuspended in 1–2 mL of sterile saline solution. Experimental details are summarized in Table 1. After the procedure, the cardiac activity of each fetus was monitored for 2 min, then re-checked 10 min later, and found to be normal in all injected fetuses.
Table 1 Experimental details of ultrasound-guided in utero injection procedures in piglets.Farrowing, clinical, and necroscopic assessmentInjections performed at GA 80 resulted in premature labor 16 days later (GA 96), leading to the delivery of 15 fetuses. All of them displayed similar developmental stages, with comparable weight and length (Supplementary Table S1); no macerated or mummified fetuses were found. These findings suggest that there was no fetal death in the hours following the procedure, but the pregnancy continued normally for 16 days until the preterm labor.
The other three sows injected at GA 108 presented a normal farrowing on GA 115, delivering a total of 37 alive piglets and 6 stillborn piglets (Supplementary Fig. S1a).
PCR analysis on DNA extracted from tail biopsies revealed that three alive piglets (#720; #725 and #779) and two stillborn piglets were GFP-positive (GFP+ Supplementary Fig. S1b), and the GFP expression in tail’s biopsies was also confirmed by visualization under a fluorescent stereomicroscopy using the GFP filter (λex = 488 nm, λem = 530 nm) (Supplementary Fig. S1c). All the GFP+ piglets, either alive or stillborn had a normal birth weight, indistinguishable from those of GFP-negative littermates (Supplementary Fig. S1d) and in line with the average weight of untreated wilde type neonatal piglets of the same strain in our facility. The two GFP+ stillborn piglets showed no signs of post-mortem or degenerative changes, such as discoloration of the skin and loss of fluids, but appeared normal, suggesting that the pigs died during the farrowing. Alive piglets (ID #720; #725 and #779) presented a regular suckling and rooting reflex; no sign of distress, lethargic phenotype, or pyrexia (Supplementary Fig. S1e) were observed compared to controls. Urine analysis revealed no alterations compared to age-matched controls (Supplementary Table S2). The #779 piglet was sacrificed 3 days after birth. During necropsy procedures, green fluorescence of the internal organs was observed (as in the stillborn piglets) when exposed to a UV lamp (Supplementary Fig. S2b). Organ weights of either alive or stillborn piglets normalised to body weight were comparable to those of uninfected, control littermates of the same age (0–3 days) (Supplementary Fig. S2c), and no macroscopic evidence of inflammatory or toxic signs was observed.
GFP+ piglets #720 and #725 were left to grow until one month of age. Their growth and behavior were comparable to un-injected siblings. No signs of distress, immune reactions (pyrexia, pain, redness, or swelling) or lethargic phenotypes were observed.
None of the sows used in the four experiments developed fever, sepsis or external inflammatory signs after the procedure. At necropsy, the uterus, liver, and lymph nodes appeared normal.
scAAV9 biodistribution and GFP expressionThe transduction efficiency was evaluated by analyzing the tissue distribution of the scAAV9 genome in fetal, neonatal, and one-month-old piglets, through PCR and digital droplet PCR (ddPCR) on DNA extracted from tissue biopsies, revealing a widespread expression of the transgene in different organs.
At the fetal stage, PCR analysis detected a GFP transgene amplicon in all tissues of fetus 1 and fetus 4 (Fig. 2a). ddPCR analysis quantified scAAV9 copies per cell in the liver (mean ± SD: 21.69 ± 25.7 vg/diploid genome), heart (0.185 ± 0.0775 vg/diploid genome), brain (0.02 ± 0.014 vg/diploid genome), muscle (0.065 ± 0.049 vg/diploid genome) and kidney (0.31 ± 0.268 vg/diploid genome) (Fig. 2b).
Fig. 2: Representative images of GFP biodistribution and expression in tissues of injected fetuses.
a PCR amplified GFP sequence was detected in different anatomical regions of fetus 1 and fetus 4. b Viral genome copies (gc) content in tissues from AAV9-treated fetuses (n = 2). c Western blotting of GFP expression in various peripheral tissues of pigs. Actin B (ACTB) served as a loading control. d Immunofluorescent staining showed that GFP was expressed in brain (cortex) and peripheral tissues of pigs after scAAV9-CAG-GFP injection. Regions are: liver (L), heart (H), cortex (C), muscle (M), kidney (K). Scalebar, 50 μm.
Accordingly, Western Blot analysis revealed a higher GFP expression in the liver, heart, and kidney, and a mild expression in skeletal muscle and brain (frontal cortex) (Fig. 2c). These results were also confirmed by immunofluorescence (Fig. 2d), suggesting that the dose of 4.5 × 1012 vg/kg conferred a good biodistribution and transgene expression at least sixteen days after injection.
In newborn and stillborn piglets, GFP was detected by standard PCR in several tissues (Fig. 3a). ddPCR was used to quantify the AAV9 copies per cell in the liver (mean ± SD: 31 ± 20 copies/cells), heart (2.66 ± 0.35 copies/cells), spleen (4.11 copies/cell), and lungs (4.44 copies/cells). AAV9 also reached the skeletal muscle, kidney, and the brain where, however, the efficiency of infection was lower (0.22 ± 0.337 copies/cell; 0.17 ± 0.136 copies/cell, and 0.113 ± 0.091copies/cell, respectively) (Fig. 3b), possibly reflecting a mixed population of AAV9-positive and -negative cells. Accordingly, Western Blot analysis showed a robust GFP expression in the liver, heart, lungs, and spleen and a moderate expression in muscle, kidney, and different areas of the brain (Fig. 3c). These results were also confirmed by immunofluorescence using an anti-GFP antibody (Fig. 3d).
Fig. 3: Representative image of GFP biodistribution and expression in different tissues of pig following transabdominal ultrasound guided injection in the fetal hearth.
a PCR amplified GFP sequence was detected in different anatomical regions. b Viral genome copies (gc) content in tissues from AAV-treated pigs (n = 3). c Western blotting of GFP expression in various peripheral tissues of pigs. Actin B served as a loading control. d Immunofluorescent staining showed that GFP was expressed in various brain regions and peripheral tissues of pigs after AAV9-GFP injection. Regions are: liver (L), heart (H), globus pallidus (GP), kidney (K), muscle (M), frontal cortex (FC), caudate (CA), thalamus (TH), cerebellum (CE), spleen (SP), lung (LU). Bars indicate the s.d. Scalebar, 50 μm.
In one-month-old piglets, ddPCR analysis showed a weak biodistribution of the transgene that was only detected in the liver of pigs #720 and #725 and in the heart of pig #725 (Supplementary Fig. S3). WB or ICC could not detect the low GFP signal in these tissues.
Assessment of anti-AAV antibody responsesOne of the major challenges in AAV-based gene therapy is the presence of circulating anti-capsid neutralizing antibodies (NAbs), which can pre-exist in patients as well as in pigs, and may prevent successful gene transfer. Moreover, high levels of circulating anti-neutralizing antibodies can develop after a single administration of gene therapy, preventing the possibility of a subsequent second treatment.
To determine the presence of pre-existing NAb to scAAV9 viral particles in our pig facility, we first measured the presence of NAb by ELISA in the serum of a sentinel sow (SOW S) and of three untreated newborn piglets. All tested animals resulted negative (Fig. 4a). Since this is a barriered facility, this result shows that there were no circulating AAVs among our animals prior to injection.
Fig. 4: Evaluation of immune response to fetal scAAV9 delivery.
a Assessment of anti-AAV antibodies responses in serum of: sentinel sow and control untreated group; scAAV9-CAG-GFP injected (GA 108) neonatal and one month old piglets and relative sows. b Quantification of cytokines fold change in scAAV9-CAG-GFP injected neonatal piglets compared to untreated neonatal controls. Results are shown as mean ± SD. P-values are indicated by *<0.05; **<0.01; ***<0.001. Blue columns indicate GFP negative piglets; green columns indicate GFP positive piglets.
To understand whether in utero administration of scAAV9-CAG-GFP triggered an immune response, we measured NAb in the serum of piglets (GFP+ and controls) and relative sows at birth and in one-month-old piglets (Fig. 4a). Also in this case, the assay found no detectable levels of NAb in any of the animals tested.
Evaluating the potential for inflammatory effects after in utero gene therapyInflammatory mediators like interferons and pro-inflammatory cytokines are frequently generated in reaction to viral infections, including those associated with AAV vectors. Understanding these immune responses is essential to evaluate the safety and efficacy of AAV-based gene therapies.
Recent preclinical studies have reported rAAV-related inflammatory toxicities in large animal models, including hepatic, cardiac, and renal toxicity, as well as neuroinflammation [24,25,26,27,28].
Because the production of inflammatory cytokines is a pathological feature of inflammation in the liver [29, 30] we have analyzed the expression of 42 cytokines in the liver of GFP+ piglets. We found a slight but significant increase only in three cytokines named: granulocyte-macrophage colony-stimulating factor (GM-CSF), growth-regulated alpha protein (GRO-α), and interferon-gamma (IFN-γ) (Fig. 4b). The upregulated levels of these cytokines may suggest the presence of an immune or inflammatory response triggered by the vector (e.g., viral capsid proteins) or by the transgene product recognized as foreign.
Histological analyses performed using hematoxylin & eosin (HE) staining did not reveal any evidence of inflammatory infiltrates or alteration to the liver’s cytoarchitecture. Furthermore, hepatocyte function remained unaltered, as demonstrated by normal periodic acid–Schiff (PAS) staining activity (Fig. 5a) observed in GFP-expressing piglets. To evaluate the possible myocardial damage associated with the AAV9-GFP injection [31] and expression, we investigated the presence of inflammatory signs or fibrosis. HE staining revealed a typical morphology of the myocardium structure without hypertrophic or degenerated cardiomyocytes. No signs of lymphocyte inflammatory infiltration were observed by acidic phosphatase (AP) staining. Further, no fibrotic areas were noted in Masson Trichrome (MT) stained sections (Fig. 5b).
Fig. 5: Histological analysis of liver and heart following fetal scAAV9 delivery.
a Histological analysis of liver morphology by HE staining and hepatocytes functionality evaluated by PAS activity; Masson Trichrome (MT) analysis did not highlight fibrotic areas in scAAV9-GFP injected fetuses. b Histological analysis of myocardium morphology by HE staining did not reveal significant alterations or inflammatory infiltrations. MT analysis did not highlight fibrotic areas in scAAV9-GFP injected fetuses. Scalebar, 100 μm.
Since recent clinical trials using rAAV have reported renal toxicity issues in addition to hepatotoxicity [32], we evaluated the kidney morphology of GFP+ and control pigs by HE staining. No obvious signs of inflammation (i.e. accumulation of white blood cells infiltrate or fibrosis) or cell death emerged (Supplementary Fig. S4), in line with normal values observed in urine analysis (Supplementary Table S2).
We then evaluated whether AAV9-GFP expression in the brain may induce neuroinflammation.
To this aim, we evaluated the distribution and the levels of ionized calcium-binding adapter molecule-1 (Iba-1) as marker of activated microglia, and glial fibrillary acidic protein (GFAP) as marker of astrogliosis, by confocal microscopy. No evident differences have emerged between GFP+ and control piglets in the frontal cortex, basal ganglia, thalamus, and cerebellum, suggesting the absence of an inflammatory response in these areas (Fig. 6).
Fig. 6: Double Immunofluorescent staining of astrocytes (GFAP in green) and microglial cells (IBA1 in red) in frontal cortex, basal ganglia, thalamus and cerebellum of scAAV9-GFP injected (#779) vs control piglet (#784).
Quantification of these parameters are shown in the graph. Data are means ± SD, unpaired t-test, NC not significative. Scalebar, 20 μm.
Comments (0)