
Remember me

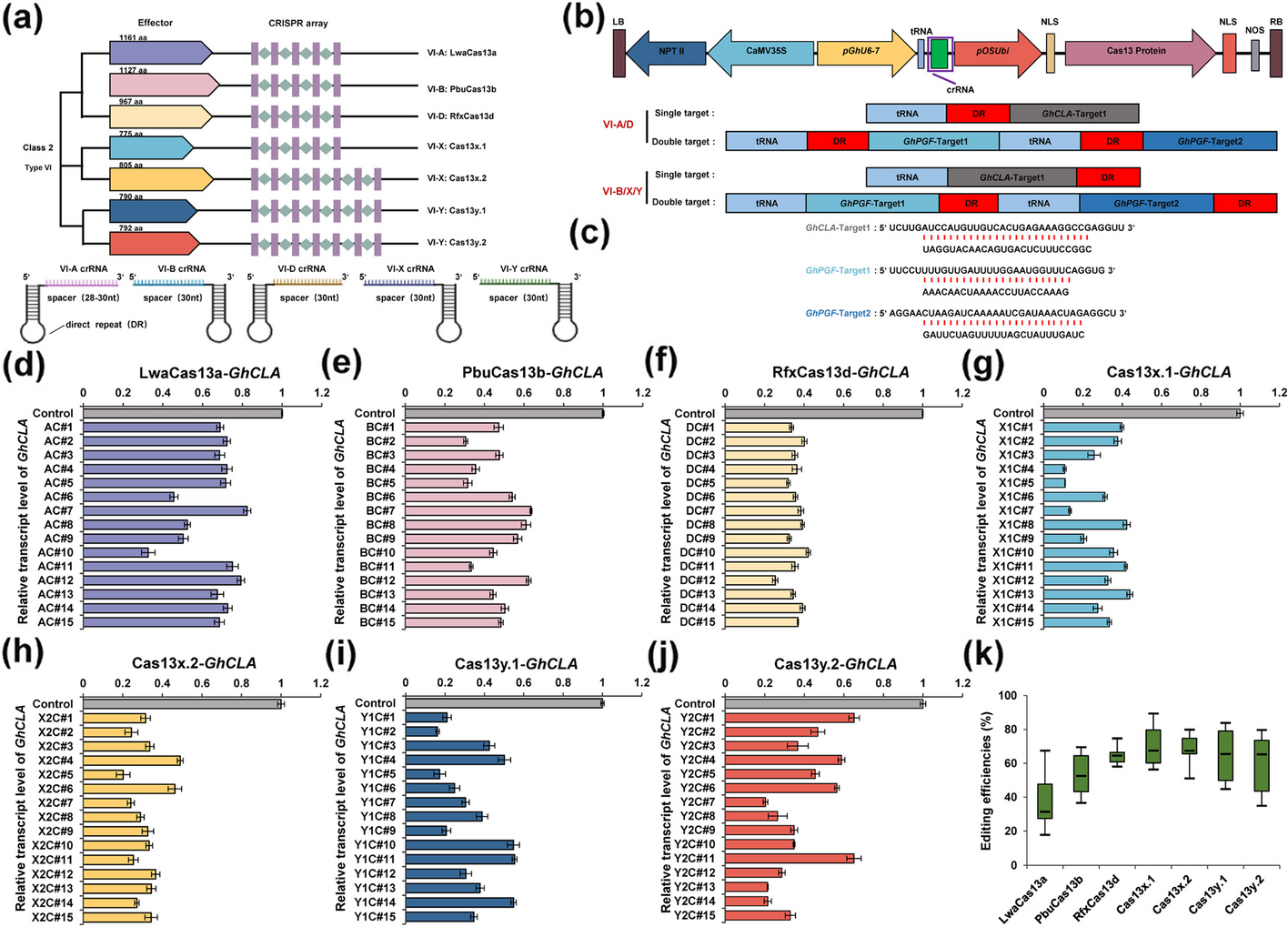
The selection of Cas13 orthologs was based on the intensity of RNA-editing activity reported both in vitro and in mammalian cells. The spatial structures of the selected proteins were predicted using AlphaFold, revealing that these proteins exhibit significantly different spatial conformations. LwaCas13a (VI-A), PbuCas13b (VI-B), and RfxCas13d (VI-D) possess more compact structures compared to Cas13y.2 (VI-Y). Interestingly, Cas13x.1 (VI-X), Cas13x.2 (VI-X), and Cas13y.1 (VI-Y) display substantial differences in their amino acid sequences, accompanied by relatively flexible spatial conformations (Additional file 1: Fig. S1a). Therefore, these Cas13 orthologs were selected for a comprehensive assessment of their editing efficiency and catalytic activities in RNA degradation within plant systems.
To engineer binary vectors for in planta expression of CRISPR/Cas13 orthologs, we codon-optimized the nucleotide sequences of LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2 based on plant genomics characteristics. Each Cas13 ortholog was co-delivered with position-matched crRNAs and their corresponding DR sequences to ensure effective targeting in plants (Fig. 1a). Cas13 orthologs were assembled using unique restriction enzymes to generate full-length clones flanked by attL1 and attL2 recombination sites. It has been demonstrated that nuclear localization signals (NLS) can significantly enhance the knockdown activity of Cas13x [22]. Therefore, two types of NLS were fused to both the C-terminus and N-terminus of the Cas13 protein. All Cas13 genes were driven by the pOSUbi10 promoter, while crRNA transcription was facilitated by the endogenous type III promoter pGhU6-7 derived from cotton, in accordance with our recent studies [47, 48] (Fig. 1b).
Fig. 1
Programmable design and efficiency assessment of various Cas13 orthologs in cotton. a Structural representations of various Cas13 orthologs and their corresponding crRNA structures. aa: amino acid. b Schematic representation of a multi-guide expression cassette design to target the endogenous transcripts GhCLA and GhPGF. c Sequence information of the target transcripts for different Cas13 proteins. d-j Transcript levels of the GhCLA gene in transgenically edited plants expressing the LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2 systems, respectively. Regenerated plants were assigned numerical identifiers based on the classification of Cas13 protein subfamilies and the target gene GhCLA. For instance, the label AC# indicates T0 generation plants that underwent LwaCas13a-mediated knockdown of the GhCLA gene, while the corresponding T1 generation is designated as AC#L. k Comparison of the knockdown efficiency of GhCLA transcripts mediated by different Cas13 orthologs in several T0 regenerated plants. Error bars represent the mean ± S.D. (n = 3)
The GhCLA (Chloroplastos alterados) gene, plays a crucial role in chloroplast development. Previous studies have shown that knockout mutants of this gene exhibit an albino phenotype [49, 50]. Additionally, silencing the GhPGF (Pigment gland formation) gene through RNAi, CRISPR/Cas9 or CRISPR/Cas12 results in a glandless phenotype [51,52,53]. In contrast to the complete knockout strategy enabled by CRISPR/Cas9, we propose that utilizing the CRISPR/Cas13-mediated knockdown approach targeting the GhCLA and GhPGF genes may yield plants with milder mutant phenotypes, such as chlorophyll loss and a reduced number of glands. Consequently, the two endogenous genes GhCLA and GhPGF were selected as target transcripts to evaluate the editing efficiency of various CRISPR/Cas13 systems. Tissue-specific expression was observed for both transcripts, revealing a statistically significant difference in RNA abundance between them (Additional file 1: Fig. S1b). A previous study indicated that the tRNA-gRNA transcription unit can enhance gRNA transcription within the CRISPR/Cas system for plant genetic editing [48, 53]. Therefore, a specific crRNA (crRNA-T) was designed to target GhCLA, leading to the construction of seven targeted vectors utilizing the tRNA-crRNA T transcription unit. In addition, two crRNAs (crRNA-T1 and crRNA-T2) were developed to target GhPGF, resulting in seven additional vectors created using the tRNA-crRNA T1-tRNA-crRNA T2 strategy (Fig. 1c & Additional file 2: Table S1). Agrobacterium-mediated transformation was applied to introduce T-DNA containing different Cas13 genes and crRNAs into the cotton plant genome (Additional file 1: Fig. S1c).
Cas13 orthologs efficiently facilitated the degradation of GhCLA transcripts using a single crRNA in cottonThrough Agrobacterium-mediated transformation, a substantial number of independently regenerated plants were obtained that harbor Cas13 systems specifically targeting the GhCLA gene using a single crRNA. For each Cas13 system, more than 15 positive T0 plants were selected to measure the transcript levels of target genes (Additional file 2: Table S2). Regenerated plants were assigned numerical identifiers according to the classification of Cas13 protein subfamilies and their target gene. For instance, the label AC# indicates T0 generation plants that underwent LwaCas13a-mediated knockdown of the GhCLA gene, while the corresponding T1 generation is designated as AC#L. Seven Cas13 genes exhibited well transcription across all T0 plants, as confirmed by quantitative reverse transcription PCR (qRT-PCR) (Additional file 1: Fig. S2). To assess the efficacy of the CRISPR/Cas13 systems in degrading targeted mRNA using a single crRNA, qRT-PCR was performed to measure the transcript levels of GhCLA. The decrease in transcript abundance is quantified as the editing efficiency of the Cas13 systems.
It was observed that the transcript levels of GhCLA were specifically downregulated in all T0 plants, while the control (non-crRNA vector) remained unchanged. The average transcript levels of GhCLA in LwaCas13a-edited plants decreased to 65.3%, with plant #10 (designated as AC#10) exhibiting the lowest recorded level at 32.7% (Fig. 1d). In contrast, the average transcript levels mediated by PbuCas13b declined to 47.4%, with the lowest level found in plant #11 (designated as BC#11), which reached 30.5% (Fig. 1e). The mRNA transcript levels were significantly downregulated in RfxCas13d-edited plants, ranging from 30 to 40%, with a notable decrease of 25.4% observed in plant #12 (designated as DC#12) (Fig. 1f). Notably, the average transcript levels of GhCLA in edited plants mediated by Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2 were measured at 29.8%, 32.1%, 35.3%, and 39.7%, respectively (Fig. 1g-j).
In conclusion, CRISPR/Cas13 systems possess the capability to target and knockdown GhCLA transcripts using a single crRNA. The average editing efficiencies were 34.7% for LwaCas13a, 52.6% for PbuCas13b, 64.3% for RfxCas13d, 70.2% for Cas13x.1, 67.9% for Cas13x.2, 64.7% for Cas13y.1, and 60.3% for Cas13y.2 (Fig. 1k). Notably, the editing efficiency of LwaCas13a has been reported to range from 58 to 88% in mammalian cells, exceeding 50% and reaching as high as 78% in rice protoplasts [25]. However, our study revealed that the editing efficiency of LwaCas13a was significantly lower in stably edited plants, with most efficiencies concentrated between 30 and 50%. A comprehensive assessment of multiple CRISPR/Cas13 nucleases confirmed that RfxCas13d, Cas13x, and Cas13y possess superior editing efficiencies along with enhanced stability, followed by PbuCas13b. In contrast, LwaCas13a exhibited the lowest level of mRNA decay activity. These findings indicate that all tested CRISPR/Cas13 orthologs are capable of performing targeted mRNA decay to modulate gene expression in cotton; however, variations in editing efficiency were observed.
Efficient degradation of GhCLA transcripts leads to a chlorophyll fading phenotype that is faithfully inherited to T1 progenySeveral independent T0 cotton plants, including AC#8 (plant #8 of LwaCas13a-GhCLA), BC#1 (plant #1 of PbuCas13b-GhCLA), DC#1 (plant #1 of RfxCas13d-GhCLA), X1C#1 (plant #1 of Cas13x.1-GhCLA), X2C#1 (plant #1 of Cas13x.2-GhCLA), Y1C#1 (plant #1 of Cas13y.1-GhCLA) and Y2C#2 (plant #2 of Cas13y.2-GhCLA), exhibited a distinct phenotype characterized by chlorophyll fading throughout the entire plant (Fig. 2a). These plants displayed varying degrees of chlorophyll fading during the genetic transformation process, which may be attributed to differences in cleavage efficiency among the various Cas13 orthologs. The chlorophyll content extracted and quantified from the leaves of T0 seedlings revealed significant reductions of 32%, 46%, 60%, 65%, 68%, 70%, and 68% for AC#8, BC#1, DC#1, X1C#1, X2C#1, Y1C#1, and Y2C#2, respectively, in comparison to control plants (Fig. 2b). This decline in chlorophyll content was consistent with the observed reduction in transcript levels of the GhCLA gene.
Fig. 2
Editing efficiency and phenotypes of various Cas13 orthologs targeting GhCLA transcripts with a single crRNA. a Chlorophyll fading phenotypes of T0 Cas13-edited plants during Agrobacterium-mediated transformation. Bars, 1 cm. b Chlorophyll content in the leaves of transgenic plants, with representative wells containing chlorophyll extract shown below (in green). Ca: chlorophyll a; Cb: chlorophyll b; Ct: total chlorophyll. Bars, 1 cm. c Transcript levels of the GhCLA gene in T1 plants expressing the LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2 systems, respectively. d Comparison of knockdown efficiency for GhCLA transcripts mediated by various Cas13 orthologs in T1 plants. e Chlorophyll fading phenotypes of T1 seedlings. Bars, 4 cm. f Quantification of chlorophyll in 4-week-old T1 seedlings, with representative wells containing chlorophyll extract shown below (in green). Error bars represent the mean ± S.D. (n = 3). Bars, 1 cm
To evaluate the potential inheritance of Cas13-mediated knockdown in the germline to T1 progeny, we assessed the editing efficiency of T1 generations. All nomenclature for the T0 and T1 generations exhibits systematic consistency; specifically, the T0 generation is designated as AC#1, while its corresponding T1 generation is labeled as AC#L1. Seedlings were cultivated in a nutrient solution until they reached the second true leaf stage. More than 10 positive T1 lines for each CRISPR/Cas13 ortholog were identified through molecular detection, confirming that Cas13 genes were well and consistently transcribed (Additional file 1: Fig. S3). An analysis of target transcripts revealed that transcript levels of the GhCLA gene were specifically downregulated in selected T1 plants, whereas the control levels remained unchanged (Fig. 2c). The average editing efficiencies were 37.3%, 44.0%, 66.0%, 68.0%, 68.4%, 68.8%, and 66.2% for LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2, respectively (Fig. 2d). These results showed a consistent trend with the editing efficiencies observed in T0 plants. Phenotypic analysis of 4-week-old T1 seedlings, along with chlorophyll measurements of the second true leaf, revealed that seedlings harboring different Cas13 orthologs exhibited varying degrees of discoloration (Fig. 2e & f). These findings suggest that RNA degradation mediated by these Cas13 systems, along with the resulting phenotypes, can be reliably inherited from T0 parental plants to their T1 progeny.
Cas13 orthologs utilize two crRNAs to effectively downregulate GhPGF transcriptsTransgenic plants expressing Cas13 orthologs targeting the GhPGF gene were generated via Agrobacterium-mediated transformation (Additional file 2: Table S3). More than 10 positive T0 plants exhibiting well-expressed Cas13 proteins for each Cas13 system were screened to evaluate editing efficiency (Additional file 1: Fig. S4). Regenerated plants were assigned numerical identifiers based on the classification of Cas13 protein subfamilies and their target gene. For instance, the label AP# indicates T0 generation plants that underwent LwaCas13a-mediated knockdown of the GhPGF gene, while the corresponding T1 generation is designated as AP#L. The results indicated that the transcript levels of GhPGF were specifically downregulated in the selected transgenic plants compared to control plants. Average decreases in GhPGF transcript levels were observed at 51.3%, 42.1%, 32.3%, 29.1%, 31.0%, 33.0%, and 34.0% for LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2, respectively (Fig. 3a-3g). Consistent with the editing effects noted on the GhCLA gene, RfxCas13d, Cas13x, and Cas13y demonstrated significantly higher efficiencies in degrading the GhPGF gene, followed by PbuCas13b and LwaCas13a (Fig. 3h). These findings further corroborate that all Cas13 systems can accurately and stably cleave targeted mRNAs using a dual-crRNA cassette approach, thereby effectively downregulating gene expression.
Fig. 3
Various Cas13 orthologs utilize two crRNAs to effectively downregulate GhPGF transcripts. a-g Transcript levels of the GhPGF gene in T0 transgenic plants expressing the LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2 systems, respectively. Error bars represent the mean ± S.D. (n = 3). Regenerated plants were assigned numerical identifiers based on the classification of Cas13 protein subfamilies and the target gene GhPGF. For instance, the label AP# indicates T0 generation plants that underwent LwaCas13a-mediated knockdown of the GhPGF gene, while the corresponding T1 generation is designated as AP#L. h Comparison of the knockdown efficiency of GhPGF transcripts mediated by various Cas13 orthologs in T0 plants. i Phenotypes of T0 transgenic plants expressing different Cas13 proteins. Bars, 5 cm. j Phenotypes of cotton bolls in T0 transgenic plants. Bars, 1 cm
Downregulation of GhPGF transcripts using CRISPR/Cas13 orthologs results in a reduced gland phenotype in cottonSeveral T0 positive plants, effectively edited with Cas13 orthologs targeting GhPGF, underwent phenotypic characterization. The total number of gossypol glands in the edited plants was reduced to varying degrees compared to control plants; however, no other significant phenotypic differences were observed (Fig. 3i). A notable reduction in the number of gossypol glands on cotton bolls was evident in the Cas13-edited plants (Fig. 3j). The number of gossypol glands on the cotton bolls decreased by 30.9%, 38.3%, 49.4%, 55.6%, 51.9%, 50.6%, and 53.1% for the plants designated as AP#3, BP#2, DP#3, X1P#2, X2P#3, Y1P#4, and Y2P#3, respectively (Fig. 4a). To more accurately assess the impact of reduced GhPGF transcripts, we quantified the gossypol content in leaves from both edited and control plants. Analysis revealed a significant reduction in gossypol levels in the leaves of T0 edited plants (Fig. 4b). As expected, the number of glands on cotton bolls and the gossypol content in the leaves were consistent with the downregulation trend of GhPGF transcripts.
Fig. 4
Downregulation of GhPGF transcripts leads to a reduced gland phenotype in cotton. a Gland number of transgenic cotton bolls per cm2. Statistical significance denoted by differing letters at P < 0.05 as determined by Student’s t-test. b Gossypol content in the leaves of T0 transgenic plants. Statistical significance denoted by differing letters at P < 0.05 as determined by Student’s t-test. c-d Transcript levels of the GhPGF gene in T1 transgenic plants expressing the LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2 systems, respectively. e Comparison of the knockdown efficiency of GhPGF transcripts mediated by various Cas13 orthologs in T1 plants. f Phenotypes of T1 seeds observed under a stereomicroscope; Control: wild-type seeds; Bars, 1000 µm. g Relative gossypol content in T1 cotton bolls. Error bars represent the mean ± S.D. (n = 3)
More than 6 independent T1 lines at the 6-week-old stage, which exhibited well-expressed Cas13 proteins, were selected for further evaluation of editing stability (Additional file 1: Fig. S5a-S5e). The transcript levels of GhPGF were consistently suppressed, demonstrating an average downregulation of 53.8% for LwaCas13a, 56.1% for PbuCas13b, 34.2% for RfxCas13d, 34.0% for Cas13x.1, 31.3% for Cas13x.2, 33.5% for Cas13y.1, and 34.7% for Cas13y.2 (Fig. 4c & d). This downregulation exhibited a similar trend with the editing efficiencies observed in T0 plants (Fig. 4e). Meanwhile, several seeds from the T1 generation were randomly selected and incubated until germination, revealing a significant reduction in gossypol glands compared to the control (Fig. 4f). Analysis of gossypol content in cotton bolls indicated a substantial decrease in T1 plants at the same developmental stage, particularly among those edited with RfxCas13d, Cas13x, and Cas13y (Fig. 4g). In the presence of CRISPR/Cas13 proteins, both the knockdown effects and the resulting phenotypes can be stably inherited across subsequent generations. Notably, T2 progeny lacking CRISPR/Cas13 proteins exhibited no significant differences in transcript levels of the targeted genes GhCLA and GhPGF compared to wild-type plants (Additional file 1: Fig. S5f).
In conclusion, all seven CRISPR/Cas13 orthologs, derived from five distinct subtypes, have been demonstrated to effectively utilize either a single crRNA or two crRNAs for the programmable knockdown of endogenous transcripts in plants. RfxCas13d, Cas13x.1, and Cas13x.2 exhibit enhanced stability and achieve editing efficiencies exceeding 58%, with a maximum efficiency recorded at 80%. Following these, Cas13y.1 and Cas13y.2 show editing efficiencies ranging from 50 to 78% and 43% to 76%, respectively. In contrast, LwaCas13a displays the lowest level of mRNA decay activity, while PbuCas13b shows slightly higher activity compared to LwaCas13a. No silencing effect on target genes was observed in T2 progeny lacking CRISPR/Cas13 proteins after the segregation of T-DNA insertion events. It has been reported that Cas13 may exert toxic effects, potentially due to its RNase activity associated with pre-crRNA processing [54,55,56,57]. However, no significant toxic effects of Cas13 have been observed in cotton embryos or mature plants, which aligns with findings from other plant studies [27, 43].
Both Cas13x.1 and Cas13y.1 orthologs mediate the simultaneous knockdown of GhCLA and GhPGF transcriptsWe integrated two guide crRNAs into a single vector to assess the capability of CRISPR/Cas13 for the simultaneous knockdown of two transcripts, aiming to extend its application for multiplexed editing in plants. The Cas13x.1 and Cas13y.1 orthologs were selected to target the GhCLA and GhPGF genes, respectively. Therefore, crRNA-T (targeting GhCLA) and the crRNA-T1 (targeting GhPGF) were chosen to construct the vectors Cas13x.1-GhCLA-GhPGF and Cas13y.1-GhCLA-GhPGF, respectively (Fig. 5a). More than 8 transgenic plants were generated through Agrobacterium-mediated transformation (Additional file 2: Table S4), in which either Cas13x.1 or Cas13y.1 protein was successfully expressed (Additional file 1: Fig. S5g & S5h). The designations X1CP# and Y1CP# refer to T0 generation plants that underwent the knockdown of both the GhCLA and GhPGF genes, mediated by either the Cas13x.1 or Cas13y.1 systems. The corresponding T1 generation plants are designated as X1CP#L and Y1CP#L.
Fig. 5
Cas13x.1 and Cas13y.1 effectively mediate the simultaneous knockdown of GhCLA and GhPGF transcripts. a Schematic representation of a multi-guide expression cassette designed to simultaneously target the endogenous transcripts GhCLA and GhPGF. b Transcript levels of the GhPGF and GhCLA genes in T0 transgenic plants expressing the Cas13x.1 system. c Transcript levels of the GhPGF and GhCLA genes in T0 transgenic plants expressing the Cas13y.1 system. b-c Error bars represent the mean ± S.D. (n = 3). The designations X1CP# and Y1CP# refer to T0 generation plants that underwent knockdown of both the GhCLA and GhPGF genes, mediated by either the Cas13x.1 or Cas13y.1 systems. The corresponding T1 generation plants are designated as X1CP#L and Y1CP#L. d Comparison of the knockdown efficiency of GhCLA and GhPGF transcripts mediated by the Cas13x.1 and Cas13y.1 systems in T0 regenerated plants. e Chlorophyll fading phenotypes of T0 transgenic plants edited with Cas13x.1 and Cas13y.1. Bars, 1 cm. f Chlorophyll content in the leaves of T0 plants expressing the Cas13x.1 and Cas13y.1 systems. Error bars represent the mean ± S.D. (n = 3). g Transcript levels of target genes in T1 transgenic plants mediated by the Cas13x.1 system. h Transcript levels of target genes in T1 transgenic plants mediated by the Cas13y.1 system. i Comparison of the knockdown efficiency of GhCLA and GhPGF transcripts mediated by the Cas13x.1 and Cas13y.1 systems in T1 plants. Error bars represent the mean ± S.D. (n = 3). j Characteristics and editing efficiencies of the seven Cas13 orthologs
The transcript levels of GhCLA and GhPGF were significantly reduced to 31.2% and 26.7%, respectively, in the X1CP#8 plant. In the Cas13x.1-GhCLA-GhPGF edited plants, the average transcription abundance of GhCLA and GhPGF transcripts decreased to 38.5% and 50%, respectively (Fig. 5b). For the Cas13y.1-GhCLA-GhPGF edited plants, transcript levels of GhCLA and GhPGF were notably diminished to 28.4% and 30% in Y1CP#2; additionally, their average transcription abundances decreased to 30.6% and 46.2%, respectively (Fig. 5c). As anticipated, Cas13x.1 and Cas13y.1 can simultaneously degrade two endogenous transcripts, resulting in a reduction in RNA abundance for both the GhCLA and GhPGF genes, with an average editing efficiency of up to 50% (Fig. 5d). These edited plants displayed a chlorophyll-fading phenotype (Fig. 5e), with chlorophyll content declining to 40.1% in X1CP#1 and 43.6% in X1CP#2, respectively. Similarly, chlorophyll content was reduced to 43.1% in Y1CP#1 and 41.1% in Y1CP#2 compared to control plants (Fig. 5f).
Six positive T1 lines for each vector were identified and selected for the analysis of RNA transcription abundance (Additional file 1: Fig. S5i). Furthermore, several seeds from T1 generation were randomly chosen and incubated until germination. The number of gossypol glands in the germinated T1 seeds showed a significant reduction compared to the control (Additional file 1: Fig. S6a). The average transcript levels of the GhCLA and GhPGF genes decreased by 60.8% and 44% for Cas13x.1 (Fig. 5g), and by 68% and 46.8% for Cas13y.1, respectively (Fig. 5h). Both Cas13x.1 and Cas13y.1 efficiently mediated multiple RNA knockdown events, achieving higher efficiency than traditional technologies (Fig. 5i). These findings provide the first evidence that CRISPR/Cas13 systems enable precise editing of multiple transcripts in plants, resulting in phenotypic traits with stable heritability. Consequently, a summary table was created to elucidate the editing characteristics associated with various Cas13 orthologs, encompassing aspects such as protein size, structural features, preferences for PAM sequences, positions of DR sequences, and editing efficiency (Fig. 5j).
Efficient RNA interference targeting of TRV transcripts in tobacco plantsSubsequently, we conducted a comprehensive evaluation of the interference activities exhibited by these CRISPR/Cas13 systems in targeting exogenous RNA viruses. TMV (Tobacco mosaic virus), one of the most destructive single-stranded RNA viruses affecting over 200 plant species, was chosen as the target virus. An analysis of the TMV genome revealed four open reading frames (ORF) that encode two replicative enzymes. Two crRNAs (crRNA1 and crRNA2) were designed to target conserved regions within TMV (Fig. 6a). TRV (Tobacco rattle virus) was selected as an additional target, and crRNA3 was designed to target the CP region of TRV RNA2 (Fig. 6b). Agrobacterium-mediated transformation was employed to generate stable transgenic plants of N. benthamiana plants expressing various Cas13 orthologs, including LwaCas13a-TMV (A-TMV), PbuCas13b-TMV (B-TMV), RfxCas13d-TMV (D-TMV), Cas13x.1-TMV (X1-TMV), Cas13x.2-TMV (X2-TMV), Cas13y.1-TMV (Y1-TMV), Cas13y.2-TMV (Y2-TMV), LwaCas13a-TRV (A-TRV), and PbuCas13b-TRV (B-TRV) (Fig. 6c & Additional file 2: Table S5). Regenerated plants were assigned numerical identifiers based on the designation of Cas13 protein subfamilies and their corresponding target viruses. For example, T0 generation plants are designated as A-TMV#, while the corresponding T1 generation plants are labeled as A-TMV#L. Notably, no abnormalities in growth or development were observed throughout the regenerative transformation process.
Fig. 6
Various Cas13 orthologs for RNA interference in tobacco plants. a Schematic representation of TMV and detailed information on the targeted sequences for various Cas13 orthologs. b Structure of TRV RNA2 along with specific details of the crRNA sequences. c Agrobacterium-mediated genetic transformation of tobacco, utilizing the K326 as the transformation recipient. Bars, 2 cm. d The TRV system expressing GFP was employed as a reporter system to evaluate Cas13 activity in transient assays. GFP: green fluorescent protein. e Monitoring GFP expression to assess Cas13-mediated viral interference activities in tobacco leaves during transient assays. Bars, 2 cm. f Relative transcript levels of different Cas13 genes in T0 transgenic tobacco plants. Regenerated plants were assigned numerical identifiers based on the classification of Cas13 protein subfamilies and their target viruses. For instance, T0 generation plants are designated as A-TMV#, while the corresponding T1 generation plants are labeled as A-TMV#L. g Disease symptoms observed in T0 transgenic plants infected with TMV at 7 dpi. Bars, 10 cm. h Virus accumulation assessed at 7 dpi using ELISA. i Virus accumulation evaluated at 7 dpi through qRT-PCR. Error bars represent the mean ± S.D. (n = 3)
An optimized TRV-RNA-based fluorescent reporter system expressing the GFP gene effectively monitors the dissemination of the CP-GFP fusion protein in tobacco leaves, thereby facilitating visualization of the viral infection process (Fig. 6d). More than 9 independent transgenic plants of A-TRV and B-TRV were selected for TRV infection, exhibiting relatively comparable levels of Cas13 protein expression (Additional file 1: Fig. S6b & S6c). Consistent with our previous findings, both Cas13 orthologs exhibited RNA interference activity against TRV. Notably, a reduction in GFP fluorescence was observed in the edited TRV-GFP leaves, with PbuCas13b demonstrating greater interference activity compared to LwaCas13a (Fig. 6e). This finding underscores the functionality and efficacy of the CRISPR/Cas13 system in mediating viral interference in planta.
Different Cas13 orthologs confer efficient interference against TMV infectionRandomly selected positive T0 transgenic plants (15 individuals per Cas13 system) were inoculated with TMV at the 3-week-old stage, and their viral resistance was subsequently assessed through symptom evaluation and molecular analysis (Fig. 6f). Wild-type tobacco plants were inoculated with TMV as positive controls, while those treated with H2O served as negative controls. We observed that TMV-infected wild-type plants exhibited typical viral symptoms at 7 days post-inoculation (dpi), including a mosaic pattern, pronounced leaf shrinkage, and curling. In contrast, T0 transgenic plants displayed no symptoms and maintained normal developmental morphology, closely resembling that of H2O-treated wild-type plants (Fig. 6g).
Serological analysis using enzyme-linked immunosorbent assays (ELISA) was performed on the selected plants. Compared to positive controls, viral titers in most transgenic plants were below detection limits (less than 0.25), indicating that the majority of tested transgenic plants exhibit resistance to TMV infection. The number of resistant plants identified by ELISA included 9 (60.0%) for A-TMV, 10 (66.7%) for B-TMV, 12 (80.0%) for D-TMV, 13 (86.7%) for X1-TMV, 13 (86.7%) for X2-TMV, 13 (86.7%) for Y1-TMV, and 13 (86.7%) for Y2-TMV (Fig. 6h). The resistance of transgenic plants to TMV was further validated through qRT-PCR assays, which revealed a significant reduction in TMV transcript accumulation in leaves at 7 dpi (Fig. 6i). These findings suggest that Cas13 systems can effectively facilitate the degradation of specific viral RNA sequences, thereby conferring resistance to TMV in transgenic plants.
Stable inheritance of resistance to TMV in transgenic tobacco progenyPositive T1 transgenic plants (10 individuals per Cas13 system) were subsequently infected with TMV to further assess their resistance at the 3-week-old stage. Wild-type plants exhibited typical mosaic leaf symptoms beginning at 7 dpi. By 40 dpi, the entire plant displayed noticeable growth deformities and dwarfism, resulting in a marked reduction in seed yield. In contrast, most transgenic lines showed only mild symptoms; notably, some transgenic plants edited with RfxCas13d, Cas13x, and Cas13y showed no apparent symptoms at 40 dpi (Fig. 7a). The markedly reduced accumulation of TMV in the leaves of transgenic plants at 40 dpi was further corroborated by ELISA and qRT-PCR assays (Fig. 7b & c). Similar results were observed in the T0 generation, suggesting that TMV resistance mediated by the CRISPR/Cas13 system is heritable.
Fig. 7
Stable inheritance of resistance to TMV in transgenic tobacco progeny. a Symptoms observed in T1 transgenic and wild-type plants infected with TMV at 40 dpi. Bars, 5 cm. b Virus accumulation was assessed at 7 dpi using ELISA in T1 transgenic plants. c Virus accumulation was quantified at 7 dpi using qRT-PCR. d Transcript levels of various Cas13 genes in T1 transgenic tobacco plants. e DAB staining was performed on infiltrated leaves of transgenic and control plants at 7 dpi. Bars, 1 cm. f H2O2 content was measured in leaves from different transgenic and wild-type plants at 7 dpi, with representative wells containing H2O2 extract shown below (in yellow). Statistical significance is denoted by differing letters at P < 0.05 using Student’s t-test. g SOD activity was measured in the leaves of various transgenic and wild-type plants at 7 dpi. Error bars represent the mean ± S.D. (n = 3). **P < 0.01
Transgenic lines exhibiting varying expression levels of Cas13/crRNA, with differences ranging from 2- to 3-fold, were selected to assess their resistance to TMV (Fig. 7d & Additional file 1: Fig. S6d). Severe mosaic symptoms were observed in TMV-infected wild-type plants; however, transgenic lines with low expression levels (e.g., A-TMV#L6) displayed mild mosaic symptoms (Additional file 1: Fig. S6e). In contrast, the transgenic lines with high expression levels (e.g., A-TMV#L2) showed no obvious symptoms. The positive correlation between Cas13/crRNA expression and the inhibition of TMV accumulation is further substantiated by the aforementioned ELISA and qRT-PCR data.
To investigate the detrimental effects after TMV infection, we further assessed the levels of reactive oxygen species (ROS), hydrogen peroxide (H2O2), and superoxide dismutase (SOD) in TMV-inoculated transgenic plants expressing various Cas13 orthologs at 7 dpi. Similarly, wild-type tobacco plants were inoculated with TMV as positive controls, while those treated with H2O served as negative controls. A higher accumulation of ROS and H2O2 was observed in TMV-infected wild-type plants, whereas transgenic plants exhibited significantly lower levels of these compounds in the infected leaves. Notably, minimal to no accumulation of ROS and H2O2 was detected in the leaves of wild-type plants treated with H2O (Fig. 7e & f). Meanwhile, the antioxidant activity of SOD was significantly elevated, peaking in wild-type plants infected with TMV, and exceeding the levels observed in transgenic plants (Fig. 7g). Collectively, these findings indicate that resistant transgenic plants exhibit minimal damage due to the effective expression of Cas13/crRNA, which facilitates the degradation of TMV transcripts and reduces the accumulation of TMV particles.
Limited off-target effects were revealed in different Cas13 transgenic cotton plantsWe further examined the specificity of Cas13 systems for transcript knockdown by assessing potential off-target effects. First, we analyzed the designed crRNAs targeting GhCLA and GhPGF transcripts for sequence similarity with other sequences using whole-transcriptome BLAST. Our findings indicated that the crRNA targeting GhCLA exhibited at least 7-nt mismatches with non-target sequences (Additional file 2: Table S6), whereas the crRNAs directed against GhPGF displayed a minimum of 5-nt mismatches with other non-target sequences (Additional file 2: Table S7). Subsequently, we performed transcriptome profiling through RNA sequencing (RNA-seq) on transgenic plants expressing various Cas13 orthologs (Additional file 1: Fig. S6f & S6g).
A comparative analysis was conducted on T1 generation plants expressing the LwaCas13a, PbuCas13b, RfxCas13d, Cas13x.1, Cas13x.2, Cas13y.1, and Cas13y.2 systems to edit the GhCLA gene. We did not observe any global downregulation of mRNA that could arise from the
Comments (0)