
Remember me

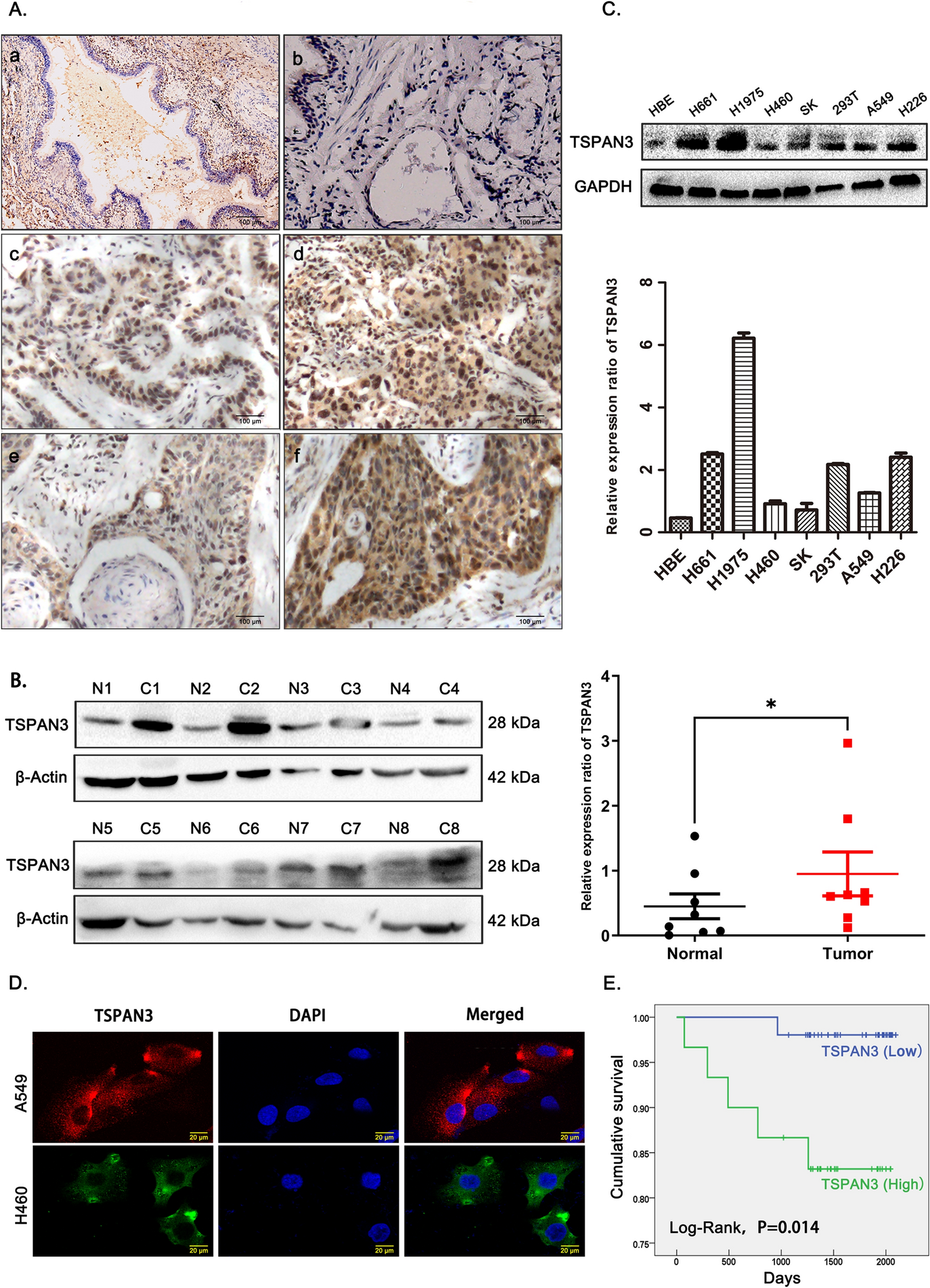



Immunohistochemistry and immunofluorescence analyses were performed to assess the expression of TSPAN3 in 81 NSCLC tissues and 2 NSCLC cell lines. TSPAN3 was found to be mainly localized in the cytoplasm (Fig. 1A, D). In addition, significantly higher levels of TSPAN3 were observed in NSCLC tissues than in the normal bronchial epithelium and submucosal glands (Fig. 1A). Western blotting was performed to assess TSPAN3 expression in eight pairs of fresh NSCLC tissues and adjacent normal tissues. NSCLC specimens were found to express prominently higher levels of TSPAN3 than their paired normal tissues (Fig. 1B). Baseline expression of TSPAN3 was also examined in six human lung cancer cell lines along with HET293T and HBE cell lines. TSPAN3 levels were consistently higher in all six lung cancer cells than those in HBE cells (Fig. 1C). Statistical analysis suggested that high TSPAN3 expression in NSCLC was positively associated with poor differentiation, lymph node involvement, and advanced pathological tumor-node-metastasis stage (P = 0.006, 0.006, 0.011, respectively; Table 1) but showed no significant association with sex, age, and histological type (P > 0.05; Table 1). Kaplan–Meier survival analysis, after following up all 81 patients for at least 3 years, further revealed that the survival period of patients with high TSPAN3 levels [IHC scores > 6, 1800.2 days, 95% confidence interval (CI) 1595.3–2005.4] was significantly shorter than that of patients with low TSPAN3 levels (IHC scores ≤ 6, 2072.8 days, 95% CI 2029.8–2115.9) (P = 0.014; Fig. 1E). Cox analyses also revealed that positive TSPAN3 expression (P = 0.043; hazard ratio of 9.139, 95% CI 1.067–78.248; Supplementary Table 2) was an independent prognostic factor in NSCLC.
Fig. 1
Expression pattern of TSPAN3 in NSCLC tissues and cell lines. A Tetraspanin 3 (TSPAN3) is expressed at low levels in normal bronchial epithelial cells (a) and submucosal gland cells (b) and expressed at high levels in NSCLC cells, well-differentiated adenocarcinoma (c), poorly differentiated adenocarcinoma (d), well-differentiated squamous carcinoma (e), and poorly differentiated squamous carcinoma (f). Magnification: × 400. B Western blot showing that TSPAN3 levels in NSCLC tissues (C) are higher than those in the surrounding normal tissues (N). β-actin served as the loading control. Relative protein expression was analyzed using ImageJ. *P < 0.05. C The endogenic TSPAN3 levels in six lung cancer cell lines, HBE cell line, and 297 T cell line as measured by western blotting and analyzed using ImageJ, GAPDH served as the loading control (n = 3 independent experiments). D Immunofluorescent probing indicates cytoplasmic localization of TSPAN3 in A549 and H460 cells. E Patients with lung cancer with high expression (IHC scores > 6) of TSPAN3 had shorter survival periods than those with low expression (IHC scores ≤ 6) of TSPAN3 (P = 0.014)
Table 1 Correlation of tetraspanin 3 (TSPAN3) expression with clinical and pathological characteristics of patients with NSCLCTSPAN3 interacts with β1 integrin via its LEL domain and upregulates β1 integrin expression in NSCLC cells.Coimmunoprecipitation was performed in A549 cells to investigate whether TSPAN3 could interact with β1 integrin. Both endogenous and transfected TSPAN3 showed efficient interaction with β1 integrin (Fig. 2A, B). As LEL domain plays a vital role in mediating the interaction of TSPAN3 with other membrane proteins, a TSPAN3 LEL-deleted construct was generated (Fig. 2C) to further explore the dependency of TSPAN3 and β1-integrin interaction on LEL domain. Coimmunoprecipitation revealed that the interaction between TSPAN3 and β1-integrin was abrogated when LEL domain was deleted (Fig. 2D). Next, to investigate the regulation of β1 integrin by TSPAN3, A549 and H460 cells (cells with moderate TSPAN3 expression) were transfected with a TSPAN3-expressing plasmid. TSPAN3 overexpression upregulated β1 integrin levels relative to the control, whereas TSPAN3 knockdown showed the opposite effect (Fig. 2E). A549 and H460 cells were also transfected with plasmids harboring the wild-type or mutant (LEL deleted; TSPAN3-ΔLEL) TSPAN3. Western blotting showed that overexpression of wild-type TSPAN3 resulted in upregulated expression of β1 integrin, which can be arrested by the deletion of LEL domain in TSPAN3-ΔLEL, further suggesting that LEL domain is required for the upregulation of β1 integrin by TSPAN3 (Fig. 2F).
Fig. 2
TSPAN3 interacts with—and upregulates—β1 integrin. A Ectopically expressed tetraspanin 3 (TSPAN3/T3) interacts with β1 integrin in A549 cells. 48 h post transfection of pCMV6-Myc-FLAG-TSPAN3 plasmid, cell lysates were immunoprecipitated with anti-FLAG antibodies or control IgG and then subjected to western blotting with anti-β1 integrin and anti-TSPAN3 antibodies. B Coimmunoprecipitation revealed that endogenous TSPAN3 interacts with β1 integrin in A549 cells. Cell lysates were immunoprecipitated with anti-TSPAN3 antibody or control IgG and then subjected to western blotting using anti-β1 integrin (ITGβ1) and anti-TSPAN3 antibodies. C Schematic of TSPAN3 LEL domain deletion. D Immunoprecipitation study indicates that TSPAN3 interacts with β1 integrin via the LEL domain. A549 cells were transfected with wild-type (WT) GFP-TSPAN3 or TSPAN3-ΔLEL mutant plasmids. After 48 h of transfection, cell lysates were immunoprecipitated with the anti-GFP antibody; the presence of β1 integrin was measured by western blotting with the anti-β1 integrin antibody. E Western blotting shows that TSPAN3 knockdown downregulated the expression of β1 integrin in A549 and H460 cells. Overexpression of TSPAN3 had the opposite effect. GAPDH served as the loading control (n = 3 independent experiments). F Overexpression of wild type TSPAN3 resulted in upregulated expression of β1 integrin, whereas overexpression of the TSPAN3-ΔLEL mutant did not exhibit any such effect. GAPDH served as the loading control (n = 3 independent experiments).G The association between TSPAN3 and β1 integrin investigated using immunohistochemistry (magnification ×200). IB, immunoblotting; IP, immunoprecipitation
We further validated the involvement of TSPAN3 in the elevated expression of β1-integrin through IHC in 20 NSCLC samples randomly selected from the 81 tissues. Correlation analysis revealed that tissues with high levels of TSPAN3 also had high levels of β1 integrin, whereas those with low levels of TSPAN3 had low levels of β1 integrin (Fig. 2G), indicating a positive correlation between the expression of TSPAN3 and β1 integrin (P = 0.022, Table 2).
Table 2 Correlation between tetraspanin 3 (TSPAN3) expression and β1 integrin levels in patients with non-small cell lung cancerTSPAN3 activates the FAK/MAPK pathway and promotes NSCLC cell proliferation via β1 integrinNext, the effects of TSPAN3 overexpression and knockdown on the phosphorylation of key FAK/MAPK pathway intermediaries downstream to β1 integrin activation [23] were investigated. Overexpression of TSPAN3 resulted in significantly increased phosphorylation of FAK, MEK, and ERK, whereas overexpression of the LEL-deleted TSPAN3 mutant did not exhibit a similar effect. This finding further demonstrates that TSPAN3 activates the FAK/MAPK signaling pathway through LEL-mediated interaction with β1 integrin. Conversely, TSPAN3 knockdown resulted in reduced phosphorylation of these key factors (Fig. 3A, B).
Fig. 3
Effect of TSPAN3 expression on the proliferation of NSCLC cell lines in vitro without the presence of FN. AWestern blot showing that overexpression of tetraspanin 3 (TSPAN3/T3) in A549 cells results in increased phosphorylation of FAK, MEK, and ERK. Knockdown of TSPAN3 in H460 cells exhibited an opposite effect. GAPDH served as the loading control (n = 3 independent experiments). B Augmented phosphorylation of FAK, MEK, and ERK, and upregulation of cyclin B1 and cyclin D1 mediated by WT TSPAN3 was abrogated by LEL domain deletion (ΔLEL). GAPDH served as the loading control (n = 3 independent experiments). C-E Colony formation, MTS assays and Edu assays showed inhibited proliferation of A549 and H460 cells in the context of TSPAN3 downregulation (n = 3 independent experiments). F–H TSPAN3 overexpression failed to promote the proliferation of A549 and H460 cells, as detected by colony formation MTS assays and Edu assays (n = 3 independent experiments). *P < 0.05; **P < 0.01; ***P < 0.001
The function of TSPAN3 in NSCLC cell proliferation was further explored. Colony formation, MTS assays, and Edu assays revealed that TSPAN3 knockdown reduced the proliferation of A549 and H460 cells (Fig. 3C–E), whereas overexpression of TSPAN3 in these cells showed negligible impact on proliferation in both cell lines (Fig. 3F–H). We speculate that this lack of progrowth effect is due to the shortage of access to integrin ligands in our in vitro culture that are otherwise highly present in vivo. Therefore, A549 and H460 cells were treated with FN (10 μg/mL) as an integrin ligand. As expected, in the presence of FN, TSPAN3 overexpression promoted the proliferation of these NSCLC cells (Fig. 4A, B). Furthermore, variations in TSPAN3 levels consistently affected the expression of proteins involved in cell proliferation. Specifically, the expression of cyclin D1 and cyclin B1 was upregulated following TSPAN3 overexpression, and overexpression of the TSPAN3-ΔLEL mutant abrogated these effects. Conversely, TSPAN3 knockdown resulted in the decreased expression of cyclin D1 and cyclin B1 (Fig. 3A, B).
Fig. 4
Effect of TSPAN3 overexpression on the proliferation of NSCLC cell lines in vitro with the presence of FN and in vivo. A, B TSPAN3 overexpression promoted the proliferation of A549 and H460 cells in presence of fibronectin (FN), as detected by colony formation, MTS and Edu assays (n = 3 independent experiments). C The volume and weight of transplanted tumors in mice injected with cells stably overexpressing TSPAN3 were greater than those in mice injected with the empty vector, whereas the transplanted tumors in mice injected with cells stably knockdown of TSPAN3 showed opposite effects [pTSPAN3 versus pControl (pCtrl), volume: 0.51 ± 0.09 cm3 versus 1.38 ± 0.13 cm3 (P < 0.01), weight: 0.80 ± 0.05 g versus 1.29 ± 0.11 g (P < 0.001); sh-TSPAN3 versus sh-control (sh-Ctrl), volume: 0.01 ± 0.01 cm3 versus 0.61 ± 0.37 cm3 (P < 0.05), weight: 0.23 ± 0.12 g versus 0.99 ± 0.11 g (P < 0.0001]]. *P < 0.05; **P < 0.01; ***P < 0.001;****P < 0.0001. FBS, bovine serum; PBS, phosphate buffered saline
The phosphorylation of FAK, MEK, ERK and level of cyclinB1, cyclinD1 increased after TSPAN3 upregulation without the presence of FN (Fig. 3A, B), which is inconsistent with the results of Colony formation, MTS assays, and Edu assays. We attribute this contradiction to the difference in time point of measurement. Increased phosphorylation of FAK, MEK, ERK and level of cyclinD1, cyclinB1 were detected at 48 h after transfection of TSPAN3 overexpression plasmid, when increased TSPAN3 caused a steady increased level of β1 integrin, resulting in the detectable transient increased in the downstream signaling pathways and cyclinD1 and cyclinB1. In contrast, the cells were continuously observed for a prolonged culture period until 144 h for colony formation, MTS assays, and Edu assays without the presence of FN, when increased level of β1 integrin no longer sustained the activation of downstream signaling pathway or levels of cyclinB1 and cyclinD1. Therefore, without FN stimulation, the enhanced proliferation in NSCLC cells was not observed in colony formation, MTS assays, and Edu assays after TSPAN3 overexpression. To prove this hypothesis, we further detected the phosphorylation of FAK, MEK, and ERK, as well as the level of cyclinD1 and cyclinB1 at multitime points after the transfection of TSPAN3 overexpression plasmid with or without FN. As the results indicated, increased level of β1 integrin was always observed through 120 h post transfection regardless of FN treatment, whereas the upregulation in phosphorylation of FAK, MEK, ERK, and the level of cyclinD1 and cyclin B1 began to attenuate at 72 h post transfection in the absence of FN. In contrast, with the stimulation of FN, the upregulation in phosphorylation of FAK, MEK, ERK, and the level of cyclinD1 and cyclin B1 was sustained until 120 h post transfection (Supplementary Fig. 1).
To further explore the effects of TSPAN3 on NSCLC proliferation in vivo, H1299 cells stably transfected with the TSPAN3-overexpressing plasmid or the sh-TSPAN3 short hairpin RNA (selected by G418, Supplementary Fig. 2) were subcutaneously delivered into nude mice for the assessment of tumor formation. Tumor volume and weight in the TSPAN3-overexpressing group were significantly higher than those in control group. In contrast, knockdown of TSPAN3 showed opposite impact (total of five nude mice per group). (Fig. 4C). All these data suggest that TSPAN3 functions as a promoter of NSCLC progression in vitro and in vivo.
TSPAN3 promotes β1 integrin recycling in NSCLC cellsTo investigate the mechanism by which TSPAN3 regulates β1 integrin production, β1 integrin (ITGB1) messenger RNA levels were first measured. PCR analysis revealed that neither TSPAN3 overexpression nor its knockdown significantly affected ITGB1 expression (Fig. 5A, B). Moreover, PCR analysis of 16 NSCLC samples showed no positive correlation between TSPAN3 and ITGB1 expression (Fig. 5C), suggesting that TSPAN3 influences β1 integrin production at the post-translational level. Considered of this finding and previous studies indicating the capacity of tetraspanins in regulating various aspects of membrane receptor trafficking, it is reasonable to hypothesize that TSPAN3 upregulates β1 integrin production via the regulation of β1 integrin trafficking and recycling. To test our hypothesis, we firstly validated the feasibility of a NZ treatment-based tracing method of integrin trafficking in our study [2, 24, 25]. Our results reveal a time sensitive, detectable restoration of the surface β1 integrin expression to baseline levels 100 min after NZ washout (Fig. 5D, E).
Fig. 5
Impact of TSPAN3 expression on the intracellular recycling of β1 integrin. A, B β1 integrin (ITGB1) mRNA levels were detected through RT-PCR in A549 and H460 cells with tetraspanin 3 (TSPAN3/T3) knockdown and overexpression (n = 3 independent experiments). C Relative mRNA expression of TSPAN3 and ITGB1 in 16 NSCLC samples as analyzed by RT-PCR. The 2−ΔΔCt value of each sample was calculated with the no. 6 sample as the reference. (P > 0.05). D Model depicting the cellular response associated with nocodazole (NZ) washout [2]. E β1 integrin expression on the surface of A549 cells at the indicated time points after NZ washout was assessed by flow cytometry. The expression values were normalized to those at 0 min (n = 3 independent experiments). F Surface expression of β1 integrin at 100 min after NZ washout relative to that at 60 min after NZ washout in A549 cells transfected with either si-TSPAN3 or si-Ctrl (n = 3). G The levels of p-FAK at different time points after NZ washout. The levels of p-FAK at 100 min after NZ washout relative to that at 60 min were calculated after normalizing to Lamin B levels (right, n = 3 independent experiments). H Immunofluorescence depicting the localization of TSPAN3 (TRITC) and β1 integrin (FITC) in A549 cells at different time points after NZ washout. Magnification ×400. *P < 0.05; **P < 0.01; ***P < 0.001
Thus, the NZ washout method was adopted to synchronize endocytosis of β1 integrins and further explore the impact of TSPAN3 on β1 integrin recycling. Flow cytometric analysis indicated that A549 cells lacking TSPAN3 were unable to restore the surface β1 integrin levels 100 min after NZ washout (Fig. 5F). Accordingly, knocking down TSPAN3 in A549 cells resulted in a failure in restoring FAK (Tyr397) phosphorylation to levels similar to those in the control group (100 min after NZ washout; Fig. 5G). Immunofluorescence on A549 cells was performed at different time points after NZ washout to verify the involvement of TSPAN3 in β1 integrin recycling. At 60 min, most of the β1 integrin had been endocytosed and transported to the perinuclear region. At this time point, most of the TSPAN3 had localized to the perinuclear region and exhibited intense colocalization with β1 integrin. At 120 min, most of the β1 integrin had returned to the membrane, and the colocalization of TSPAN3 and β1 integrin was lost (Fig. 5H). Taken together, these data suggest that TSPAN3 is involved in, and promotes, the recycling of β1 integrin.
TSPAN3 promotes β1 integrin recycling by facilitating its sorting into Rab11a recycling endosomesTo further explore the specific mechanism underlying the involvement of TSPAN3 in β1 integrin recycling, mass spectrometric analysis was performed. Consistent with reports that the Rab11-dependent mechanism is one of the major routes for integrin recycling, the mass spectrometric data (Supplementary Figs. 3 and 4) indicated that TSPAN3 could promote β1 integrin recycling via Rab11a. The interaction between TSPAN3 and Rab11a was further verified in A549 cells through coimmunoprecipitation (Fig. 6A, B). Immunofluorescence also revealed the colocalization of TSPAN3 and Rab11a in the cytoplasm (Fig. 6C). Furthermore, this interaction did not depend on the LEL domain of TSPAN3 (Fig. 6D). Interestingly, a dynamic change in the colocalization of TSPAN3 and Rab11a was observed at different time points after NZ washout. At 60 min post NZ washout, TSPAN3 also colocalized with Rab11a, which showed an overlapping pattern with that of the dynamic change of colocalization between TSPAN3 and β1 integrin after NZ washout. The colocalization was maintained up until 120 min after NZ washout (Fig. 6E), Additionally, TSPAN3 knockdown or overexpression did not affect Rab11a levels (Supplementary Fig. 5). Combining previous studies and the present findings, we inferred that TSPAN3 may interact with Rab11a and β1 integrin, thereby facilitating β1 integrin sorting into Rab11a recycling endosomes.
Fig. 6
Effect of TSPAN3 on β1 integrin recycling. A Coimmunoprecipitation assays showing the interaction between exogenous tetraspanin 3 (TSPAN3/T3) and endogenous Rab11a in A549 cells. A549 cells were transfected with pCMV6-Myc-FLAG-TSPAN3 constructs. After 48 h of transfection, cell lysates were immunoprecipitated with anti-FLAG antibodies or control IgG and subjected to western blotting with anti-Rab11a and anti-FLAG antibodies. B Coimmunoprecipitation assay showing the interaction between endogenous TSPAN3 and Rab11a in A549 cells C Immunofluorescence showing the localization of TSPAN3 (TRITC) with Rab11a (FITC) in the cytoplasm of A549 cells. Magnification ×400. D Western blot indicating that the interaction between TSPAN3 and Rab11a does not depend on the LEL domain of TSPAN3. After 48 h of transfection for wild-type GFP-TSPAN3 or TSPAN3 mutant, cell lysates were immunoprecipitated with the anti-GFP antibody; the presence of Rab11a in the anti-GFP-precipitated complex was examined through western blotting with the anti-Rab11a antibody. E Immunofluorescence showing the colocalization of TSPAN3 (TRITC) and Rab11a (FITC) in A549 cells at different time points after nocodazole (NZ) washout (magnification ×400). F Pearson correlation coefficient of the colocalization between Rab11a and β1 integrin at 60 min after NZ washout. G Immunofluorescence showed that TSPAN3 knockdown (si-T3) resulted in reduced colocalization of β1 integrin and Rab11a (60 min after NZ washout) and the surface expression of β1 integrin (120 min, indicated by arrowheads). Magnification ×400. H Immunofluorescence showed that TSPAN3 knockdown (si-T3) resulted in increased colocalization of β1 integrin and LAMP1 at random time without NZ washout. Magnification ×400. I Pearson correlation coefficient of the colocalization between LAMP1 and β1 integrin at random time without NZ washout. J Western blot indicating that Rab11a knockdown attenuated the TSPAN3-induced increase in β1 integrin, p-FAK (Tyr397), p-MEK, p-ERK, cyclin B1, and cyclin D1 expression. GAPDH served as the loading control (n = 3 independent experiments). K Schematic of the mechanism of TSPAN3 regulating β1 integrin intracellular recycling, TSPAN3 promotes β1 integrin intracellular recycling by facilitating β1 integrin sorting into Rab11a recycling endosomes, which reduces β1 integrin lysosomal degradation and enables more rapid resurface of integrin
Immunofluorescence on A549 cells at different time points after NZ washout was performed next to explore the changes in the interaction between β1 integrin and Rab11a after TSPAN3 knockdown. The results showed that at 60 min after NZ washout, most of the β1 integrin had been endocytosed in both the si-TSPAN3 and control groups. However, in the control group, the endocytosed β1 integrin showed significant colocalization with Rab11a, whereas the colocalization between β1 integrin and Rab11a was lower after TSPAN3 knockdown (Fig. 6F, G). Correspondingly, at 120 min after NZ washout, most of the β1 integrin in the control group had returned to the periphery of the cell, whereas most of the β1 integrin in the si-TSPAN3 group remained in the cytoplasm (Fig. 6G). After the knockdown of TSPAN3 in A549, increased colocalization of β1 integrin and LAMP-1(the biomarker of lysosome) were found at all time points (Fig. 6H, I). Moreover, immunoblotting showed that si-Rab11a transfection of both A549 and H460 cells attenuated the TSPAN3-induced elevation in β1 integrin, p-FAK, p-MEK, p-ERK, cyclin D1, and cyclin B1 levels (Fig. 6J), which further supported our hypothesis. Based on all above-mentioned findings, we concluded that TSPAN3 promotes β1 integrin recycling, upregulates β1 integrin levels, and promotes NSCLC cell proliferation through a Rab11a-dependent mechanism (Fig. 6K).
Comments (0)