
Remember me


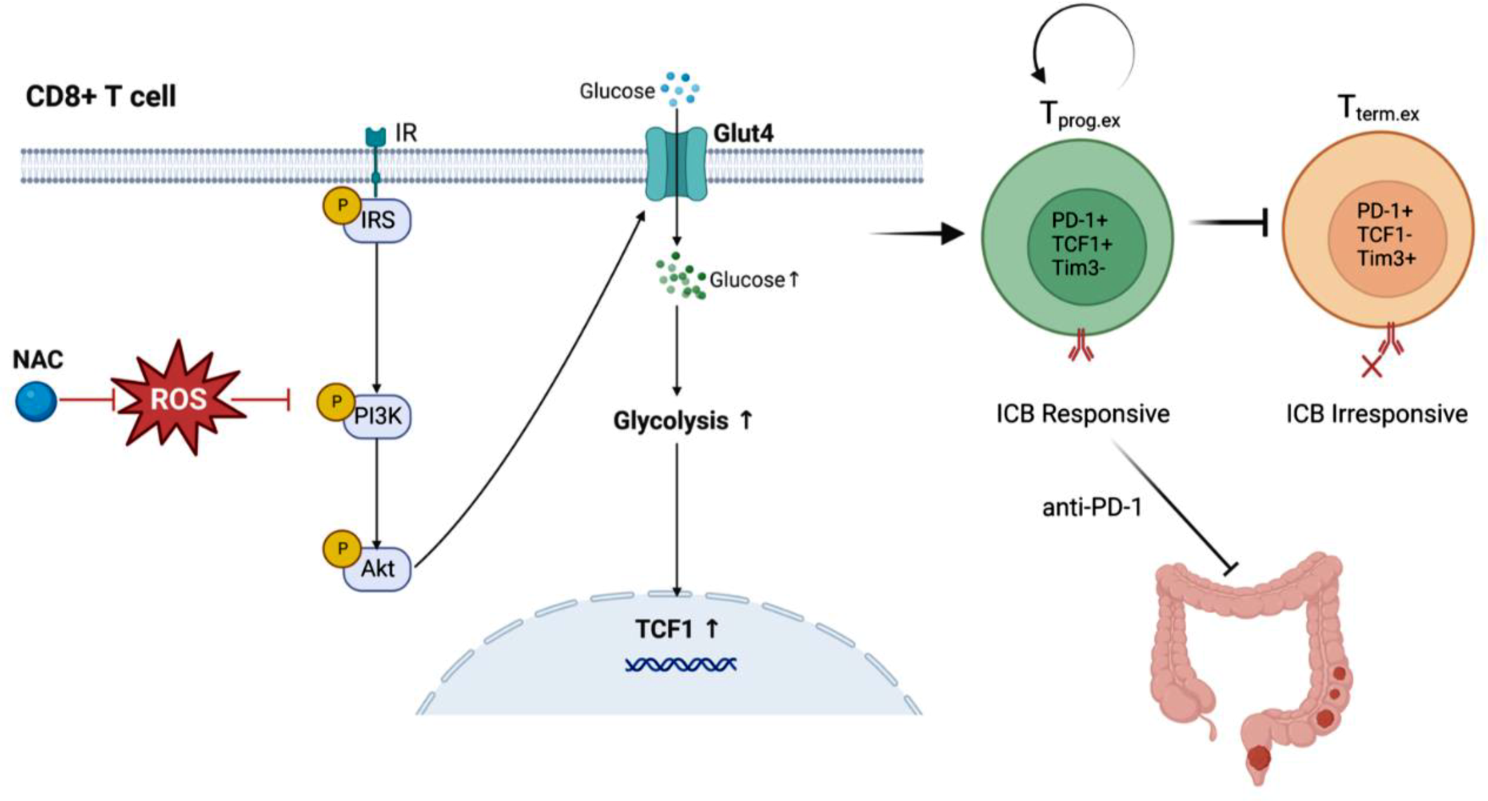
In this study, we examined the impact of NAC on the proliferation of in situ implanted MC38 cells in mice. To replicate the clinical course, established tumors underwent treatments within a specified period (NAC for days 5–20 and anti-PD-1 for days 10, 14, and 18) (Fig. 1A). The NAC group had a smaller tumor volume and lower weight compared to the control group (Additional File 1A, B).
Fig. 1
NAC enhances the effect of PD-1 antibody against colorectal cancer progression. (A) Schematic representation of the treatment of tumor-bearing mice with NAC and/or PD-1 antibodies. Tumor-engrafted mice were randomized into four groups and treated with NAC (days 5–21; n = 5 mice), anti-PD-1 (days 10, 14, and 18; n = 5 mice), their combination (n = 5 mice), or vehicle (control; n = 5 mice). Body weight changes during the experiment (B). Tumor volumes (left) and tumor weight (right) at day 21 are shown (C, D). Tumor growth of MC38 cells implanted subcutaneously in syngeneic mice treated as in (E) (days 5–21; n = 5 mice). Tumor size changes during the experiment (F). Tumor volumes (left) and tumor weight (right) at day 21 are shown in (G). We harvested tumors on day 21 post-transplantation and analyzed the cell number of the infiltrated CD4+ and CD8+ cells (H, I). We used 10 mice in both the control group and the NAC group in (H, I), so there were 10 mice in both groups in the tumor tissue analysis. As the number of cells in the mesenteric lymph nodes of some mice after experimental treatment could not meet the requirements for flow analysis, we had to discard the data of these mice, resulting in 6 mice in each group in SLNs analysis. Data are expressed as means ± standard error of the mean. ANOVA (analysis of variance) test for (B, D, F, G and I ); *P < 0.05, **P < 0.01, ***P < 0.005
Subsequently, we examined the impact of integrating NAC with the anti-PD-1 monoclonal antibody, RMP1-14, on the proliferation of in situ implanted tumors. The combination of NAC with PD-1 blockade had no significant effect on the body weight of mice (Fig. 1B). The combined treatment resulted in decreased tumor growth compared to the individual efficacy of each treatment (Fig. 1C, D). We obtained similar conclusions in the mouse subcutaneous tumor experiments (Fig. 1E–G).
To investigate the influence of NAC on tumor antigen-specific T cells, we conducted flow cytometry experiments on the 21st day after implantation, using tumor tissues and mesenteric lymph nodes from tumor-bearing mice. Our findings indicated no statistically significant disparity in the proportion of CD4+ cells to total CD45+ cells between the two groups in tumor tissues and sentinel lymph nodes (SLNs). However, the NAC group exhibited a higher ratio of CD8+ cells to CD45+ cells (Fig. 1H, I). We also found that the ratio of CD8+ cells to CD45+ cells in the combined group was higher than that in the PD1 group in tumor tissues and SNLs (Additional File 1D). Additionally, the NAC group had a greater ratio of PD1+ cells to CD45+CD8+ cells in tumor tissues (Additional File 1 C). These findings suggest that NAC can raise the amount of CD8+ T cells specific to tumor antigens and the expression of PD-1 in CD8+ T cells, potentially hindering the progression of CRC.
NAC-induced sensitivity to PD-1 blockade is mediated by an antitumor CD8+ T cell responseThe effects of NAC (0, 1, 5, 10, 15, and 20 mM) on the proliferation of human-derived CRC cell lines (HCT116 and HT29) and murine-derived CRC cell lines (MC38 and CT26) were observed using CCK8 and clone formation assays. Based on the MC38 cell colony-formation assay, the number of colonies formed in the 1 mM NAC group was not statistically different from that in the control group; the number of colonies formed in the 5 and 10 mM groups was higher than that in the control group, and that in the 15 and 20 mM NAC groups was lower than that in the control group (Fig. 2A, B). Based on MC38 cells and the CCK8 assay, on day 5, the OD value of the 1 mM NAC group was higher than that of the control group; the OD of the 5, 10, and 15 mM NAC groups was not statistically different from that of the control group; and the OD of the 20 mM NAC group was lower than that of the control group (Fig. 2C). Other CRC cell lines exhibited comparable results. The results suggested that 1–10 mM NAC did not inhibit the proliferation of CRC cells, while 15 and 20 mM NAC had significant cytotoxicity.
Fig. 2
CD8+ T cells are required for NAC-induced sensitivity to anti-PD-1 blockade. (A) Colon cancer lineage cells are cultured under normal or gradient concentrations of NAC conditions for about 14 days. The experiment was repeated three times with similar results. The statistical plot of the number of colony formations in Fig. 2B. Data are expressed as three technical replicates. (C) CT26, MC38, HCT116, and HT29 cells were cultured in 0, 1, 5-, 10-, 15-, and 20-mM NAC medium for 1–5 days, and then CCK8 reagent was incubated for 1 h—statistical plots of OD values at 450 nm. (D) Schematic of the experiment in which MC38 cells cultured under normal or STS-like conditions for 48 h were inoculated subcutaneously in the left flank of syngeneic mice. Seven days after injection, MC38 cells cultured under normal conditions were subcutaneously inoculated in the right flank of these mice, and animals were treated (or not) with anti-PD-1 (days 10, 14, and 18 after the last MC38 inoculation; n = 5 mice per group). (E) Left subcutaneous tumor picture at the top, and the right subcutaneous tumor picture at the bottom. (F) Left flank tumor size follow-up (left) and tumor size at day 21 (right). The number of tumors per group was 5: normal precondition (precond); normal precond + anti-PD-1; NAC precond; NAC precond + anti-PD-1. (G) Right flank tumor size follow-up (left) and tumor size on day 21 (right). (H) Tumor-bearing mice were treated with the NAC + anti-PD-1 regimen as indicated, in the presence or absence of depleting antibodies (days 5, 10 and 15). The number of mice per group was 5: control; NAC + anti-PD-1; NAC + anti-PD-1 + anti-CD4; and NAC + anti-PD-1 + anti-CD8. (I) Tumor volumes of these mice at day 21 (right) are shown. Data are expressed as means ± standard error of the mean. ANOVA test for (B, C, F, G, and I); *P < 0.05, **P < 0.01, ***P < 0.005
According to a previous study [30], we evaluated whether NAC treatment increased the immunogenicity of MC38 cells. We preconditioned MC38 cells and subcutaneously inoculated the mice with normally cultured MC38 cells on the right flank (naïve tumor side) and treated them with anti-PD-1 or vehicle, as outlined in Fig. 2D. No significant difference in tumor volume was observed on the immunized side after the PD-1 blockade. No significant abscopal effect associated with anti-PD-1 treatment was observed on the naïve tumor side (Fig. 2E–G), indicating that NAC did not exert anti-tumor effects by enhancing the immunogenicity of MC38 cells.
According to a previous study [30], some conditions can increase the immunogenicity of cancer cells through inducing autophagy; therefore, we tested the autophagy marker LC3-II/LC3-I ratio by western blotting (WB). Additionally, we explored whether NAC could exert anti-tumor effects by inhibiting the expression of PD-L1. WB experiments were carried out on MC38 cells pretreated with a medium containing 10 mM NAC for 48 h, as well as on tumor tissues from mouse in situ tumors. The results showed that NAC did not affect the expression of PD-L1 or the LC3-II/LC3-I ratio in MC38 cells in vivo (Additional File 2 A) or in vitro (Additional File 2B).
Concerning the identity of effector T cells, the anti-tumor efficacy of NAC + anti-PD-1 treatment in MC38 tumors was entirely nullified by the depletion of CD8+ T cells. Depletion of CD4+ T cells resulted in a mild suppression of the anti-tumor effect (Fig. 2H, I). These findings suggest that the susceptibility of mouse CRC tumors to PD-1 blockade, induced by NAC, is influenced by the presence of cytotoxic CD8+ cells.
Effect of NAC on apoptosis, proliferation, and secretory function of CD8+ T cellsMouse CD8+ T cells were sorted and cultured in normal RPMI 1640 medium or RPMI 1640 medium containing 10 mM NAC for 5 days, and then the cells were used for the cell flow assay. The results showed that the proportion of annexin+ cells in the NAC group was lower than that in the control group (Fig. 3A), suggesting that NAC reduced the apoptosis of CD8+ T cells. The number of Ki67+ and CD25+ cells increased in the NAC group (Fig. 3B, C), suggesting enhanced cell proliferation and activation. Microscopic images and cell counting results (Additional File 3A) showed that the NAC group had a higher cell density.
Fig. 3
Effect of NAC (10 mM) on the proliferation, activation, and secretion of CD8+ T cells in vitro. (A) CD8+ T cells were sorted from mouse spleens and cultured under normal or 10 mM NAC conditions for 5 days. Representative flow cytometry plots showing the expression of Annexin and PI among CD8+ T cells. (B, C). Flow cytometry plots and quantification showing the expression of Ki67 and CD25 among CD8+ T cells. (D). Flow cytometry plots and quantification showing the expression of Granzyme B, TNF-α, and IFN-γ after cell activation cocktail stimulation. (E) Cell supernatants at 5 days of culture were taken and subjected to Luminex liquid suspension microarray experiments. Cytokine secretion from CD8+ T cells was detected using the Bio-Plex Pro Mouse Cytokine Grp I Panel 23-plex kit. (F) Supernatants of CD8+ T cells at 5 and 7 days of culture were taken and analyzed for IL-2, Granzyme B, TNF-α, and IFN-γ concentrations using ELISA assay. P-values are from two-tailed unpaired t-tests (A, B, C, D, F); *P < 0.05, **P < 0.01, ***P < 0.005
The proportion of Granzyme B+ and IFN-γ+ cells in CD8+ T cells in the NAC group was lower than in the control group, while the difference in the proportion of TNF-α+ cells was not statistically significant (Fig. 3D). Luminex liquid-phase suspension microarray experiments were conducted to detect cytokine secretion by CD8+ T cells using a Bio-Plex Pro Mouse Cytokine Grp I Panel 23-plex kit (#M60009RDPD, Bio-Rad, USA). The results showed that the concentration of IL-2 was higher in the NAC group than in the control group, while the cytotoxic factors TNF-α and IFN-γ were significantly lower in the NAC group than in the control group (Fig. 3E). To further determine the effect of NAC on the secretory function of CD8+ T cells, the concentrations of IL-2, Granzyme B, TNF-α, and IFN-γ in cell supernatants at 5 and 7 days of culture were measured using ELISAs (Fig. 3F). The results showed that the concentration of IL-2 in the NAC group was higher than that in the control group, and the concentrations of Granzyme B, TNF-α, and IFN-γ in the NAC group were lower than those in the control group.
To clarify the effect of NAC on the secretion function of CD8+ T cells in vivo, mouse tumor tissues were taken for cell flow analysis on the 21st day of in-situ colorectal tumor implantation. Cytometry results showed there was no significant difference in the proportion of Granzyme B+ and IFN-γ+ cells in CD45+CD8+ cells between the NAC group and the control group (Additional File 3B). Peripheral blood of the same mice was detected by Luminex liquid suspension chip, and the results showed no significant difference in the levels of TNF-α, IFN-γ, and IL-2 between the NAC group and the control group (Additional File 3 C).
NAC promotes TCF1+PD1+CD8+ T cell differentiation and reduces Tterm.ex. DifferentiationTCF1+PD1+CD8+ T cells were considered as Tprog.ex. and PD1+Tim3+CD8+ T-cells as Tterm.ex [6]. To explore the effect of NAC on CD8+ T cell differentiation, CD8+ T cells from mice cultured for 5 days were used for cell flow cytometry. In the NAC group, the proportion of TCF1+CD8+ T cells was higher than that in the control group, as were the proportions of TCF1+PD1+CD8+ T cells (Fig. 4A, B), suggesting that NAC could induce Tprog.ex. differentiation. The proportion of PD1+Tim3+CD8+ T cells in the NAC group was lower than that in the control group (Fig. 4C), suggesting that NAC inhibits Tterm.ex. formation. WB showed that TCF1 expression increased in CD8+ T cells in the NAC group (Fig. 4D, Additional File 4A), confirming that NAC can promote TCF1 expression in CD8+ T cells.
Fig. 4
NAC promotes TCF1+ PD1+ CD8+ T cell differentiation and reduces Tterm.ex. differentiation. (A) CD8+ T cells were sorted from mouse spleens and cultured under normal or 10 mM NAC conditions for 5 days. Representative flow cytometry plots showing the expression of TCF1 among control and NAC group CD8+ T cells. (B, C) Flow cytometry plots and quantification showing the expression of TCF1+PD1+CD8+ cells (B) and the frequencies of PD1+Tim3+ Tterm.ex. (C). (D) CD8+ T cells cultured for 5 days were taken for protein extraction, and TCF1 protein expression level was detected by WB assay. (E–G) On day 21 of in situ colorectal implant tumors, mouse tumor tissues were harvested for cell flow experiments. First, cells were gated on CD45+CD8+ cells. Then, flow cytometry plots and quantification show the frequencies of CD44+CD62L+ cells (E), TCF1+PD1+ cells (F), and PD1+Tim3+CD8+ T cells (G). P-values are from two-tailed unpaired t-tests (B, C) and ANOVA (analysis of variance) test (E–G); *P < 0.05, **P < 0.01, ***P < 0.005
To investigate the effect of NAC on the differentiation of tumor antigen-specific CD8+ T cells, tumor tissues of mice were collected on day 21 of the in situ tumor implantation for cell flow experiments (Additional File 4B), which showed that the difference in the CD44+CD62L+CD8+ T cell (Tm) ratio between the four groups in tumor tissues and SLNs was not statistically significant; the ratio of TCF1+PD1+CD8+ T cells in the NAC group was higher than that in the control group and anti-PD1 group (Fig. 4E). The difference in the TCF1+PD1+CD8+ T cell ratio between the NAC + anti-PD1 and NAC groups was not statistically significant, and the proportion of PD1+Tim3+CD8+ T cells in the NAC group was lower than that in the control and anti-PD1 groups (Fig. 4F). The difference in the proportions of PD1+Tim3+CD8+ T cells between the NAC + anti-PD-1 and NAC groups was not statistically significant (Fig. 4G). These results suggest that NAC promotes CRC-associated PD1+TCF1+CD8+ T cell differentiation and reduces the formation of Tterm.ex. but has no significant effect on Tm. Flow cytometry analysis of mouse mesenteric lymph nodes was performed, and similar results were obtained (Additional File 2 C).
NAC enhances glucose uptake and glycolysis of CD8+ T cellsTo explore the potential molecular mechanisms by which NAC affects CD8+ T cell differentiation, CD8+ T cells were cultured in a normal 1640 medium and NAC-containing medium for 5 days. Transcriptome sequencing analysis showed that Slc2a4, which encodes Glut4, was significantly upregulated in the NAC group compared to the control group (Fig. 5A). PCR results showed that the mRNA levels of Slc2a1, Slc2a3, and Slc2a4 in CD8+ T cells in the NAC group were higher than those in the control group, with Sl2a4 being the most significant (Fig. 5B). WB results showed that Glut4 protein levels in the NAC group were higher than those in the control group (Fig. 5C, Additional File 5B). These results confirmed that NAC enhanced Glut4 mRNA and protein expression levels in CD8+ T cells.
Fig. 5
NAC promotes elevated Glut4 expression and glucose metabolism in CD8+ T cells. (A) CD8+ T cells were sorted from mouse spleens and cultured under normal or 10 mM NAC conditions for 5 days. Two groups of CD8+ T cells were then taken and analyzed by transcriptome sequencing. (B) According to transcriptome sequencing results, NAC increased Slc2a4 mRNA levels in CD8+ T cells. Two groups of CD8+ T cells, which were also cultured in vitro for 5 days, were taken, and the mRNA expression levels of Slc2a1,2,3,4,5 of the two groups were examined by PCR assay (Slc2a2 was under-expressed). Data are expressed as ratios relative to β-actin. (C) WB assay was performed to detect the expression of Glut4 in CD8+ T cells of both groups. (D, E) CD8+ T cells from control and NAC groups cultured in vitro were taken for Seahorse assay to detect cellular OCR (D) and ECAR (E) levels. (F–H) Control and NAC CD8+ T cells cultured in vitro were taken, and intracellular glucose concentration, ATP concentration, and lactate concentration were detected by glucose uptake assay (D) and ATP (D), and lactate concentration detection assays (D). (I) CD8+ T cells cultured in vitro for 5 days were taken, and then the relative mRNA levels of PFKM, PKLR, HK3, LDHB, and LDHD were detected by PCR. Data are expressed as ratios relative to β-actin. P-values are from two-tailed unpaired t-tests (B–H); *P < 0.05, **P < 0.01, ***P < 0.005
Seahorse tests showed that the ECAR of cells in the NAC group was higher than that in the control group (Fig. 5E), and the difference in OCR between the two groups was not statistically significant (Fig. 5D), suggesting that NAC could promote glycolysis in CD8+ T cells. The glucose uptake assay showed that cellular 2-Deoxyglucose 6 phosphate (2-DG6P) content in the NAC group was higher than that in the control group, suggesting that NAC promoted glucose uptake by CD8+ T cells (Fig. 5F). ATP and lactate concentration assays showed that the ATP and lactate concentrations in the CD8+ T cells of the NAC group were higher than those in the control group (Fig. 5G, H), supporting that NAC could promote glycolysis.
In vitro transcriptome sequencing results were screened for key enzymes in the glycolytic pathway with significant differences in expression: PFKM, PKLR, HK3, LDHB, and LDHD. The mRNA levels of these enzymes were detected by PCR. The results showed that the mRNA levels of PFKM, HK3, LDHB, and LDHD were higher in the NAC group than in the control group and that there was no significant difference in the mRNA levels of PKLR between the two groups (Fig. 5I), further confirming that NAC promoted CD8+ T cell glycolysis. Furthermore, NAC increased the proportion of Glut4+CD8+ cells in mouse TILs (Fig. 6A), suggesting that NAC increases Glut4 expression in mouse CRC tumor antigen-specific CD8+ T cells. Immunofluorescence analysis of tumor tissues further confirmed the increased Glut4 expression in CD8 + T cells in the NAC group (Fig. 6B).
Fig. 6
NAC promotes Glut4 expression on colorectal cancer tumor associated CD8+ T cells. (A) Mouse in situ colorectal cancer was modeled, and tumor tissue was taken from mice on day 21 for flow cytometry. Flow cytometry plots and quantification showing the expression of Glut4+ cells (gated on CD45+CD8+ cells). (B) Mouse colorectal cancer tumor tissues were taken and subjected to immunofluorescence staining for DAPI (blue), CD8+ (red), and Glut4 (green). (C, D) MC38 cells were cultured with normal or 10 mM NAC medium for 48 h. Then, MC38 cells were taken for WB assay to detect the expression level of Glut4 (C). Immunofluorescence staining was performed to detect the expression level of Glut4 (D). (E) An in situ colorectal cancer model was constructed in mice. Vehicle or NAC gavage were given until 21 days after implantation. Tumor tissues from both groups were taken for WB assay to detect the expression level of Glut4. P-values are from two-tailed unpaired t-tests (A, B, C, D); *P < 0.05, **P < 0.01, ***P < 0.005
We investigated the effect of NAC on Glut4 in MC38 cells. The results of WB and immunofluorescence showed that NAC had no significant effect on Glut4 expression in MC38 cells in vitro (Fig. 6C, D) or in vivo (Fig. 6E, F). This suggests that the effect of NAC on Glut4 in CD8+ T cells has some specificity. The statistical results and an image of the tumor are shown in Additional File 5 A, C,D.
NAC regulates glucose metabolism and differentiation of CD8+ T cells via the IR/PI3K/Akt pathwayWe hypothesized that NAC could affect Glut4 expression by regulating the IR/PI3K/Akt pathway through reactive oxygen species (ROS) and affect CD8+ T cell differentiation through Glut4-regulated glucose metabolism.
Using cytometry, we demonstrated that NAC reduced ROS and mitochondrial reactive oxygen species (mtROS) levels in mouse CD8+ T cells in vitro (Additional File 6 A, B), and ROS levels in mouse tumor tissues in vivo (Additional File 6D). In addition, NAC reduced mitochondrial depolarization in CD8+ T cells in vitro (Additional File 6 C). Proteins were extracted from CD8+ T cells cultured for 5 days, and the protein levels of key molecules in the IR/PI3K/Akt pathway were examined by WB. The results showed that the expression levels of p-IRS1 (Tyr895), PI3K, p-PI3K (Tyr458), Akt, and p-Akt (Ser473) were higher in the NAC group than in the control group (Fig. 7A).
Fig. 7
NAC regulates glucose metabolism by activating the IR/PI3K/Akt pathway in CD8+ T cells. (A) CD8+ T cells cultured for 5 days were taken for protein extraction, and the expression levels of Akt, p-Akt (Ser473), PI3K, p-PI3K (Tyr458), IRS1, p-IRS1 (Tyr895) were examined by WB assay. (B, C) CD8+ T cells were cultured in normal medium and NAC medium for 3 days, and the cells were grouped into Ctrl, NAC, Akt1-IN (Inhibitor abbreviated as IN), and NAC + Akt1-IN groups. Ctrl and NAC groups were rehydrated with the normal medium or medium containing 10 mM NAC; the Akt-IN group was rehydrated in the normal medium with MK-2206 (7.5 µM), and the NAC + Akt-IN group was rehydrated in NAC medium with MK-2206 (7.5 µM). Then, after 48 h of further culture, CD8+ T cells were collected for WB assay to detect Glut4 expression. (C). (D) CD8+ T cells cultured in vitro were taken and divided into four groups, as shown in the method above: Ctrl, NAC, Akt1-IN, and NAC + Akt1-IN. The ECAR levels of cells in the four groups were detected by Seahorse assay. (E–G) Intracellular glucose concentration, ATP concentration, and lactate concentration were detected by glucose uptake assay (E) and ATP (F) lactate concentration detection assays (G). (H–J) Flow cytometry quantification show the frequencies of PI+Annexin+ cells (H), Ki67+ cells (I), and TCF1+PD1+ cells (J) among CD8+ T cells. Plots were shown in Additional File 5. P-values are from two-tailed unpaired t-tests (A) and ANOVA (analysis of variance) test (C–G); *P < 0.05, **P < 0.01, ***P < 0.005
To clarify whether NAC regulates Glut4 expression through the IR/PI3K/Akt pathway, considering Akt is the core molecule of this pathway, validation experiments were performed using the Akt inhibitor, MK-2206 Dihydrochloride. WB results showed that Glut4 expression in the NAC + Akt-IN group was lower than that in the NAC group (Fig. 7B). PCR results showed that Slc2a4 mRNA levels in the NAC + Akt-IN group were lower than those in the NAC group (Fig. 7C). These results suggested that Akt inhibitors antagonized the effect of NAC on mRNA and protein levels of Glut4 in CD8+ T cells.
The seahorse assay showed that the ECAR levels of cells in the NAC + Akt-IN group were lower than those in the NAC group (Fig. 7D). The glucose uptake assay showed that the cellular 2-DG6P level was significantly lower in the NAC + Akt-IN group than in the NAC group (Fig. 7E). ATP and lactate concentration assays showed that the intracellular ATP and lactate concentrations were significantly lower in the NAC + Akt-IN group than in the NAC group (Fig. 7F, G). These results suggest that the Akt inhibitor antagonizes the effect of NAC to enhance glycolysis and glucose uptake in CD8+ T cells.
To determine whether inhibition of Akt could antagonize the effect of NAC on apoptosis, proliferation, and differentiation of CD8+ T cells, after 3 days of culture, CD8+ T cells were transferred to a new corresponding cell culture medium with MK-2206 (7.5 µM), and the culture was continued for 48 h more. The CD8+ cells were then taken for cell flow experiments. The proportion of apoptotic CD8+ T cells in the NAC + Akt-IN group was higher than that in the NAC group (Fig. 7H, Additional File 7 A), and the proportion of TCF1+PD1+CD8+ cells and Ki67+CD8+ T cells was lower than that in the NAC group (Fig. 7I, J, Additional File 7B, C). Akt inhibitors effectively antagonize the anti-apoptotic, pro-proliferative, and pro-TCF1+PD1+ subtype differentiation effects of NAC on CD8+ T cells.
NAC enhances glucose metabolic differentiation of CD8+ T cells via Glut4WB analysis showed that Glut4 expression in the NAC + shGlut4 group was lower than that in the NAC + Vector group (Fig. 8A), suggesting that Glut4 shRNA could antagonize the effect of NAC to elevate Glut4 protein expression in CD8+ T cells.
Fig. 8
NAC regulates glucose metabolism by Glut4 in CD8+ T cells. CD8+ T cells were cultured in vitro and divided into Ctrl group, Ctrl + vector group, NAC + vector group, and NAC + shGlut4 group, cultured in 1640 medium and 1640 medium containing 10 mM NAC concentration for 48 h, respectively. The Ctrl + vector and NAC + vector groups were transfected with a medium containing empty virus, and the NAC + shRNA group was transfected with a medium containing Slc2a4 shRNA lentivirus. After 24–48 h, they were replaced with the corresponding virus-free culture medium, and cells were collected for the experiment after another 24 h culture. (A) The expression levels of Glut4 were examined by WB assay (relative to actin). (B) The ECAR levels of cells in the four groups were detected by Seahorse assay. (C–E) Intracellular glucose concentration, ATP concentration, and lactate concentration were detected by glucose uptake assay (C) and ATP (D) lactate concentration detection assays (e). (F–H) Flow cytometry plots and quantification show the frequencies of PI+Annexin+ cells (F), Ki67+ cells (G), and TCF1+PD1+ cells (H) among CD8+ T cells. P-values are from ANOVA (analysis of variance) test (A–H); *P < 0.05, **P < 0.01, ***P < 0.005
The Seahorse assay showed that the ECAR level of cells in the NAC + shGlut4 group was significantly lower than that in the NAC + Vectro group (Fig. 8B). The glucose uptake assay showed that the cellular 2-DG6P content in the NAC + shGlut4 group was significantly lower than that in the NAC + Vectro group (Fig. 8C). The ATP and lactate concentrations in the NAC + shGlut4 group were significantly lower than those in the NAC + Vectro group (Fig. 8D, E). These results suggest that Glut4 shRNA can antagonize the effect of NAC in enhancing glycolysis and glucose uptake by CD8+ T cells.
Cell flow analysis revealed that the proportion of apoptotic CD8+ T cells was higher in the NAC + shRNA group than in the NAC group (Fig. 8F, Additional File 8 A), the proportion of TCF1+PD1+CD8+ cells was lower than that in the NAC group (Fig. 8G, Additional File 8B), and the proportion of Ki67+CD8+ T cells was lower than that in the NAC group (Fig. 8H, Additional File 8 C). This suggests that Glut shRNA antagonizes the anti-apoptotic and pro-proliferative effects and induction of TCF1+PD1+ subtype differentiation by NAC in CD8+ T cells.
Comments (0)