
Remember me


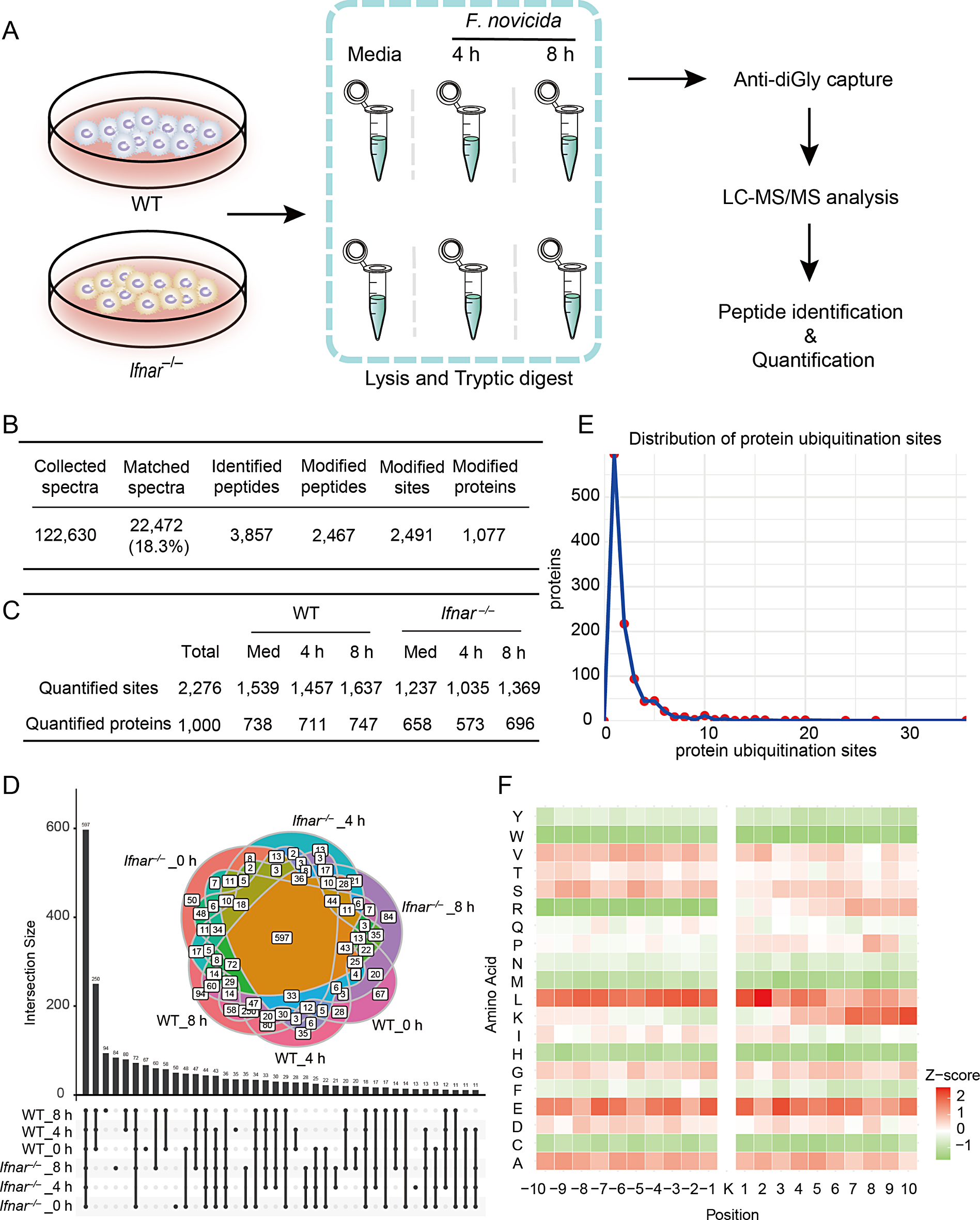
To examine the role of IFN-I in the cellular ubiquitinome in the response of BMDMs to F. novicida infection, we performed diGly proteomics for quantitative ubiquitination site analysis in both WT and interferon-alpha receptor (IFNAR)-deficient (Ifnar–/–) BMDMs with and without F. novicida infection (Fig. 1A). This approach integrates stable isotope labeling in cell culture (SILAC) with immunoprecipitation enrichment of diGly-terminated peptides following trypsin digestion of ubiquitinated proteins. The dynamics of the host ubiquitinome induced by F. novicida infection were investigated at 4 and 8 h post infection (Fig. 1A). Mass spectrometry analysis yielded 122,630 secondary spectra, with 22,472 spectra successfully matched, leading to the identification of 3,857 peptides, of which 2,467 peptides were ubiquitinated. A total of 1,077 proteins with 2,491 ubiquitination sites were identified (Fig. 1B, Table S1). Quantitative analysis revealed differences in the ubiquitination profiles between WT and Ifnar–/– BMDMs and a decrease in the number of quantified ubiquitination sites in Ifnar–/– BMDMs compared with WT BMDMs (Fig. 1C). To verify the ubiquitination sites detected in our data, we conducted literature search for some well-known proteins and confirmed 25 ubiquitination sites in 23 proteins reported in previous studies (Table S2). These data indicate the potential loss of original ubiquitination sites on certain proteins in Ifnar–/– BMDMs and reveal a dynamic change in ubiquitin modification sites after F. novicida infection.
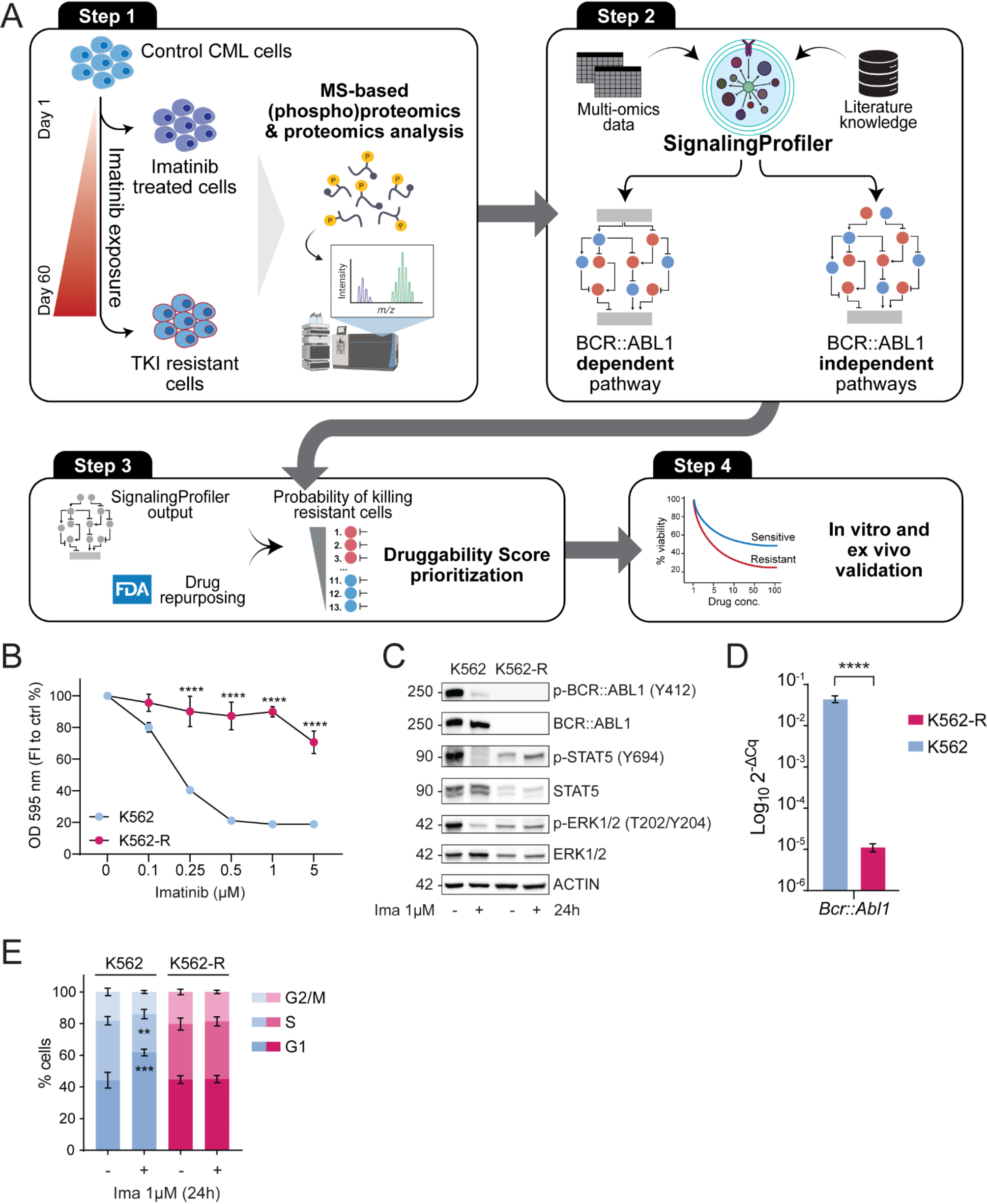
Fig. 1
Ubiquitin landscape in BMDMs post infection with F. novicida. A, Workflow of diGly capture and LC‒MS/MS analysis of uninfected and F. novicida (200 MOI)-infected WT and Ifnar–/– BMDMs for 4 and 8 h. B, Total identified peptides, ubiquitination sites, ubiquitinated peptides and proteins identified in both the WT and Ifnar–/– BMDMs in (A). C, Comparison of quantified ubiquitination sites and ubiquitinated proteins between the WT and Ifnar–/– BMDMs in (A). D, Venn diagram analysis of shared and specific ubiquitinated proteins in the WT and Ifnar–/– BMDMs in (A). E, Distribution analysis of the number of quantified ubiquitination sites per protein. F, Heatmap showing the motif enrichment analysis of upstream and downstream amino acids of all identified modification sites. Red indicates that this amino acid is significantly enriched close to the modification site, and green indicates that this amino acid is significantly reduced close to the modification site
We subsequently performed Venn diagram analysis to compare the quantification of ubiquitination sites in different samples, including shared and specific ubiquitination sites, in WT and Ifnar–/– BMDMs at both 4 and 8 h post infection (Fig. 1D). Notably, 597 ubiquitination sites were commonly detected in all the samples. Sixty-seven, 35, and 94 ubiquitination sites were specifically detected in WT BMDMs at 0, 4, and 8 h after F. novicida infection, respectively. In contrast, 50, 13, and 84 specific ubiquitination sites were detected in Ifnar–/– BMDMs at 0, 4, and 8 h, respectively (Fig. 1D). The distribution analysis of ubiquitination sites across the identified proteins revealed that the majority of proteins contained one or two ubiquitination sites, suggesting that the ubiquitination sites were conserved among these samples (Fig. 1E). Motif analysis of the amino acids upstream and downstream of the ubiquitination sites revealed an increased frequency of leucine (L) and glutamic acid (E) residues surrounding the ubiquitination sites (Fig. 1F). Collectively, these data reveal a dynamic ubiquitin landscape during F. novicida infection and a differential ubiquitination profile between WT and Ifnar–/– BMDMs, suggesting the subtle role of ubiquitination regulated by IFN-I in regulating the interplay between host cells and bacterial pathogens.
Profile of changes in the ubiquitination site in WT BMDMs during F. novicida infectionTo investigate the changes in ubiquitination triggered by F. novicida, we quantified the dynamic profile of total ubiquitinated sites within each protein in WT BMDMs in response to F. novicida infection. We found that the total ubiquitination of 454 proteins was reduced by F. novicida infection; F. novicida infection caused increased ubiquitination of 194 proteins, and the ubiquitination of 263 proteins was comparable before and after F. novicida infection (Fig. 2A). The top list of proteins with reduced ubiquitination included the cell integrity and cytoskeleton-regulating proteins VIM and TUBA1B, the ribosomal subunit proteins RPS3 and RPS27A, the nuclear envelope protein LMNA, the receptor for colony stimulating factor CSF1R, the ATPase VCP, and the ubiquitin-regulating proteins USP5, USP25, and RNF213 (Fig. 2B). The greatest increase in protein ubiquitination was observed for the cell death-modulating protein PARP14, the serine proteinase inhibitor SERPINB6, the NADPH oxidase complex subunit CYBB, the actin-binding protein gelsolin GSN, the adaptor protein STIP1, the ion channel protein KCNAB2, the phosphofructokinase PFKL, the GTPase GVIN1, the translin-associated factor X TSNAX, and the transcription factor NME2 (Fig. 2B). KEGG enrichment analysis revealed that the proteins involved in pathways related to cell death, protein degradation, and inflammatory responses were mostly ubiquitinated during F. novicida infection (Fig. 2C). The Venn diagram showed the results of the statistical analysis of proteins with increased, decreased, and comparable degrees of ubiquitination involved in the cell death, protein degradation, and inflammatory response pathways. Approximately two-thirds of the proteins in these pathways exhibited dynamic changes in ubiquitination during F. novicida infection (Fig. 2D). Furthermore, many proteins involved in regulating pyroptosis, apoptosis, necroptosis, and autophagy, such as NLRP3, IL-1β, GBP2, IL-1α, RIPK1, and BID, exhibited dynamic changes in ubiquitination in response to F. novicida infection (Fig. 2E). Taken together, these results indicate that host cells coordinate multiple signaling pathways through ubiquitination and underscore the important role of dynamic ubiquitination in regulating immune responses and pathogenicity during F. novicida infection.
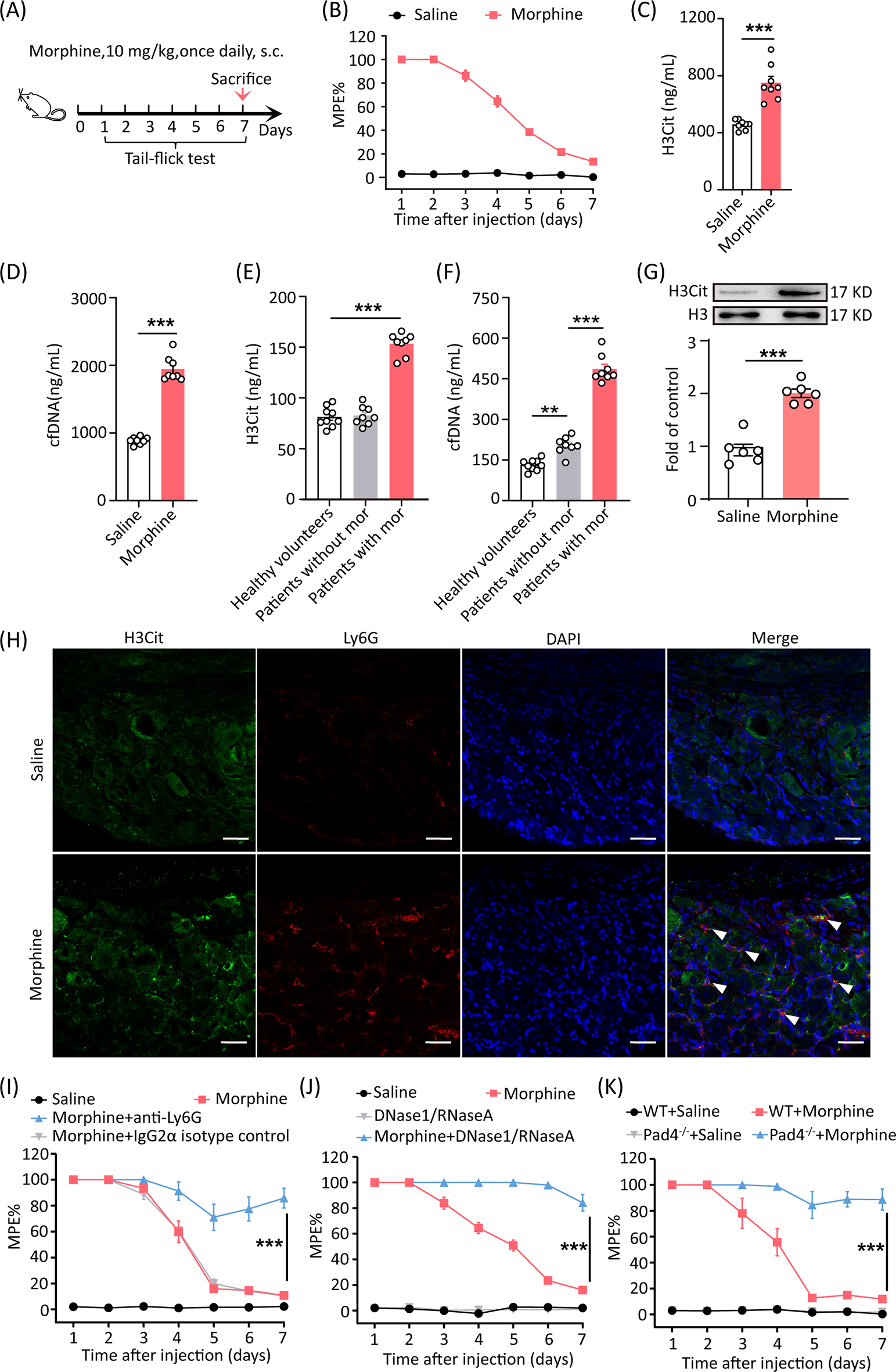
Fig. 2
Analysis of ubiquitination dynamics in WT BMDMs during F. novicida infection
A, Quantitative analysis of changes in the ubiquitination of WT BMDM proteins in response to F. novicida (200 MOI) infection for 4 and 8 h. Ubi_up indicates proteins with increased ubiquitination in F. novicida-infected WT BMDMs. Ubi_down indicates proteins with reduced ubiquitination in F. novicida-infected WT BMDMs. Comparable indicates proteins with similar levels of ubiquitination between uninfected and F. novicida-infected WT BMDMs. B, Heatmap showing the top ten proteins with the greatest decrease (Ubi_down) and increase (Ubi_up) in ubiquitination in WT BMDMs after F. novicida infection (200 MOI) for 4 and 8 h. C, KEGG enrichment analysis of increased and decreased ubiquitinated proteins in WT BMDMs after F. novicida infection. D, Venn diagram analysis of the proteins with decreased (Ubi_down), increased (Ubi_up), and comparable (Ubi_others) ubiquitination after F. novicida infection, which are involved in the pathways of cell death, protein degradation, and the inflammatory response. E, Heatmap showing ubiquitinated proteins involved in the pyroptosis, apoptosis, necroptosis, and autophagy pathways. Ubi_down indicates decreased ubiquitination after F. novicida infection; Ubi_up indicates increased ubiquitination after F. novicida infection; Ubi_others indicates comparable ubiquitination between uninfected and F. novicida-infected WT BMDMs
Effects of IFN-I signaling on ubiquitination in BMDMsIFN-I plays both positive and negative roles in host defense against extracellular and intracellular bacterial infection [23, 24]. To examine the role of IFN-I in host protein ubiquitination during F. novicida infection, we compared the changes in the ubiquitination of host proteins between WT and Ifnar–/– BMDMs at 0, 4, and 8 h after F. novicida infection. Ubiquitination sites in 502 proteins were reduced in Ifnar–/– BMDMs compared with WT BMDMs (50%), ubiquitination sites in 160 proteins were increased in Ifnar–/– BMDMs (16%), and ubiquitination sites in 336 proteins were comparable between WT and Ifnar–/– BMDMs (34%) (Fig. 3A, B). The similar trend of dynamic change in the ubiquitination proteins involved in inflammatory response, protein degradation, and cell death were observed in WT and Ifnar–/– BMDMs during F. novicida infection (Fig. 3C). The proteins whose ubiquitination was increased by F. novicida infection in WT BMDMs further increased ubiquitination in Ifnar–/– BMDMs, and vice versa (Fig. 3A, C), suggesting that IFN-I signaling was essential for both the increase and decrease of ubiquitination in response to F. novicida infection. In the subsequent analysis of the quantified proteins, we identified 37 E3 ubiquitin ligases, of which 6 E3 ligases exhibiting increased ubiquitination and 15 E3 ligases showing reduced ubiquitination in Ifnar–/– BMDMs compared with WT BMDMs. In addition, 14 deubiquitinating enzymes were detected, of which 3 proteins showing increased ubiquitination and 7 proteins showing reduced ubiquitination in Ifnar–/– BMDMs compared with WT BMDMs (Fig. 3D, Table S3). The decrease in total ubiquitination, K48- and K63-linked ubiquitination in WT and Ifnar–/– BMDMs induced by F. novicida infection was confirmed by western blot analysis (Fig. 3E, S1A). To elucidate the specific impact of IFNAR deficiency on ubiquitination in BMDMs during F. novicida infection, we conducted a functional enrichment analysis of proteins according to the altered ubiquitination levels. These analyses included Gene Ontology (GO) classification and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis and aimed to reveal the biological significance of alterations in ubiquitination in the absence of IFNAR. Increased ubiquitination in Ifnar–/– BMDMs was observed for proteins predominantly enriched in biological processes such as membrane proteins, organelle fusion, and ubiquitin-dependent protein degradation (Fig. 3F). Additionally, these proteins were enriched in pathways involving SNARE interactions with vesicular transport, small cell lung cancer-related pathways, protein processing in the endoplasmic reticulum, and the NF-κB signaling pathway according to KEGG analysis (Fig. S1B). In contrast, proteins with decreased ubiquitination in Ifnar–/– BMDMs were significantly enriched in biological processes related to cytoplasmic translation, the regulation of protein metabolism, cytoskeletal organization, and the regulation of innate immune responses (Fig. 3G). Furthermore, the proteins associated with decreased ubiquitination in Ifnar–/– BMDMs were enriched in critical metabolic pathways, such as spinocerebellar ataxia, ribosomal structure, proteasome, prion disease, and peroxisome pathways, according to KEGG analysis (Fig. S1C).
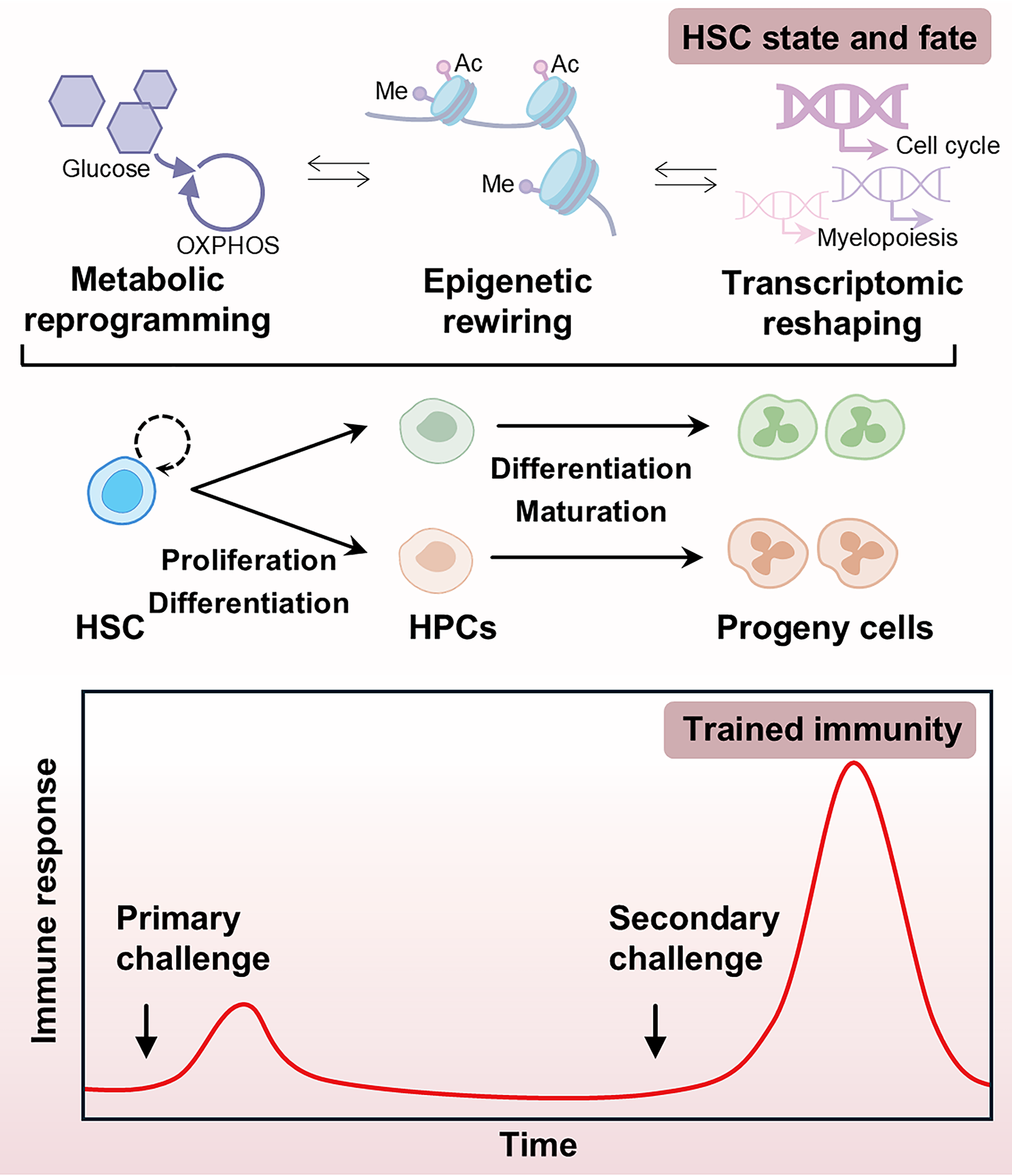
Fig. 3
Impact of IFNAR deficiency on protein ubiquitination and function. A, Plot showing the dynamic changes in protein ubiquitination levels between WT and Ifnar–/– BMDMs before and after F. novicida (200 MOI) infection for 4 and 8 h. The x-axis indicates 0, 4, and 8 h post infection. The y-axis indicates the relative level of protein ubiquitination, and the level of ubiquitination in WT BMDMs at 0 h was normalized to 1. Down indicates decreased ubiquitination in Ifnar–/– BMDMs compared with WT BMDMs; Up indicates increased ubiquitination in Ifnar–/– BMDMs compared with WT BMDMs; Comparable indicates comparable ubiquitination between WT and Ifnar–/– BMDMs. B, Quantification of ubiquitinated proteins in Ifnar–/– BMDMs compared with WT BMDMs. Ubi_down indicates decreased ubiquitination in Ifnar–/– BMDMs compared with WT BMDMs under both uninfected and F. novicida-infected conditions; Ubi_up indicates increased ubiquitination in Ifnar–/– BMDMs compared with WT BMDMs under both uninfected and F. novicida-infected conditions; Comparable indicates comparable ubiquitination between WT and Ifnar–/– BMDMs under both uninfected and F. novicida-infected conditions. C, Heatmap showing the ubiquitination levels of representative proteins involved in inflammatory response, protein degradation, and cell death pathways in WT and Ifnar–/– BMDMs at 0, 4, and 8 h after F. novicida infection. D, Venn diagram showing the overlap of proteins with increased (Ubi_up) and decreased (Ubi_down) ubiquitination in Ifnar–/– BMDMs compared with WT BMDMs of E3 ubiquitin ligases and deubiquitinating enzymes with and without F. novicida infection. E, Immunoblot analysis of total ubiquitin (anti-Ub, P4D1) in WT and Ifnar–/– BMDMs without infection (Med) or infected with F. novicida (200 MOI) for 4 and 8 h. F, G,Gene Ontology (GO) enrichment analysis of the differentially ubiquitinated proteins in Ifnar–/– BMDMs compared with WT BMDMs. Plot showing the enriched biological process (BP), molecular function (MF), and cellular component (CC) terms of the increased ubiquitination group (D) and decreased ubiquitination group (E) in Ifnar–/– BMDMs compared with WT BMDMs under both uninfected and F. novicida-infected conditions. Data are representative of 3 independent experiments with similar results for (E)
IFN-I signaling affects the ubiquitination of proteins regulating cell death and ubiquitination pathways in response to F. novicida infectionTo specifically elucidate the effect of IFN-I on the ubiquitination of proteins during F. novicida infection, we profiled the top list of decreased and increased ubiquitination sites of proteins in Ifnar–/– BMDMs compared with WT BMDMs (Fig. 4A), as well as the dynamic changes at each specific ubiquitination site in these proteins (Fig. S2). Notably, the ubiquitination sites were highly detected in glycolysis-related proteins, including glucose-6-phosphate isomerase (GPI), transketolase, an enzyme involved in the pentose phosphate pathway (TKT), aldolase A, a crucial glycolytic enzyme (ALDOA), poly (ADP‒ribose) polymerase member 14 (PARP14), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), in WT BMDMs but not in Ifnar–/– BMDMs (Fig. 4A, S2). In addition, the actin cytoskeleton-regulated proteins MYH9 and WDR1, the ribosomal protein RPL4, the E3 ubiquitin ligase ring finger protein RNF213, and the guanylate binding protein GBP2 were identified as ubiquitination site-rich proteins in WT BMDMs; however, these ubiquitination sites were not detected in Ifnar–/– BMDMs (Fig. 4A, S2). On the other hand, the inflammatory cytokine IL-1β, membrane transporters, receptors such as TNFRSF1B, VAMP8, ATP7A, FUNDC2, and the voltage-gated proton channel HVCN1 were highly ubiquitinated in Ifnar–/– BMDMs but not in WT BMDMs (Fig. 4A, S2). Furthermore, the dynamic ubiquitination changes of NLRP3 and GAPDH in response to F. novicida infection were confirmed by western blot analysis (Fig. S3A-C).
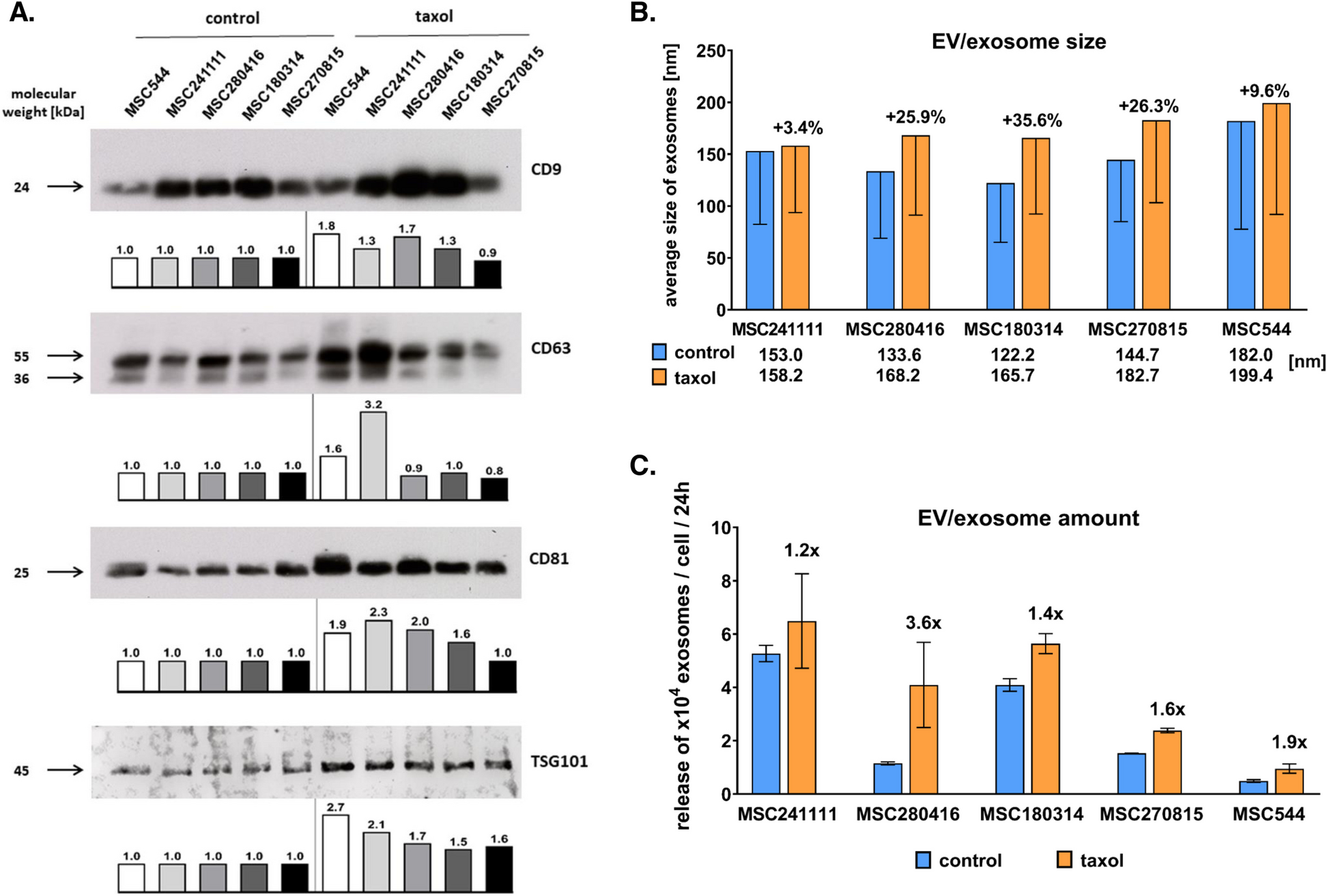
Fig. 4
Characterization of the proteins with the most increased and decreased ubiquitination sites in WT BMDMs compared with those in Ifnar–/–BMDMs. A, Heatmap showing the ubiquitination levels of the proteins with the greatest decrease and increase of ubiquitination in WT BMDMs compared with those in Ifnar–/– BMDMs with and without F. novicida infection. B, C, Predicted protein structure of PARP14 (B) and MYH9 (C) labeled with the total identified ubiquitination sites (red dots) in both WT and Ifnar–/– BMDMs. The yellow and blue frame-labeled regions are shown with enlarged images. D, E, Predicted protein structures of RNF213 (D), GBP2, ALDOA, and TKT (E) labeled with the total identified ubiquitination sites (red dots) in both WT and Ifnar–/– BMDMs
We further investigated the ubiquitination sites within different proteins closely. In PARP14 and MYH9, the ubiquitination sites were mostly identified within the conserved macro- and PARP domains of PARP14 and the myosin motor and coiled coil domains of MYH9 (Fig. 4B and C). Instead, the ubiquitination sites observed in the RNF213, GBP2, ALDOA, and TKT proteins were not restricted to conserved domains (Fig. 4D and E).
Interaction network and dynamics of major ubiquitinated proteins in cell death pathwaysCell death plays essential roles in both host defense against bacterial infection and bacterial evasion of host immune responses [25]. In this study, we found that ubiquitinated proteins associated with cellular death pathways such as pyroptosis, necroptosis, and apoptosis were significantly enriched in both WT and Ifnar–/– BMDMs (Fig. 5A). Subcellular localization analyses of these proteins revealed a predominant distribution in the cytoplasm (45.76%) and nuclear localization (21.47%). A notable fraction localized to the cell membrane (10.17%) and the mitochondrial environment (7.91%) (Fig. 5B). Further investigation into the ubiquitination patterns of proteins involved in cell death pathways confirmed that IFNAR deficiency caused mostly decreased ubiquitination levels of the proteins involved in the cell death pathways (Fig. S4).
Fig. 5
Network analysis of ubiquitinated proteins involved in cell death pathways. A, Enrichment analysis of all ubiquitinated proteins quantified in WT and Ifnar–/– BMDMs that are involved in cell death pathways, including pyroptosis, apoptosis, necroptosis, cellular senescence, and autophagy pathways. B, Subcellular localization analysis of all ubiquitinated proteins identified in WT and Ifnar–/– BMDMs that are involved in cell death pathways. C, Interaction network analysis of ubiquitinated proteins involved in cell death pathways, highlighting the key hub proteins ranked by betweenness centrality (BC). Proteins located close to the center are considered hub proteins within the network. D, Heatmap showing the dynamic ubiquitination changes at specific sites of central hub proteins within the protein interaction network of the cell death pathways
Given the impact of IFN-I on ubiquitination dynamics, we further analyzed the protein interaction networks within cell death pathways affected by ubiquitin modifications. The proinflammatory cytokine IL-1β, the tyrosine kinase JAK1, the apoptotic regulator BCL2L1, the multifunctional scaffolding protein p62/SQSTM1, and the molecular chaperone HSP90AB1 are central hub proteins in these networks, potentially playing a leading role in signal transduction and crosstalk during F. novicida infection (Fig. 5C). The resulting heatmap accurately displays the levels of ubiquitination at specific sites on five central hub proteins within the protein interaction network of the cell death pathway. Notably, there was a significant increase in the level of ubiquitination at 5 sites of IL-1β and 10 sites of SQSTM1 in the absence of IFNAR (Fig. 5D). In contrast, the ubiquitination levels of the HSP90AB1 protein significantly decreased at 14 sites in Ifnar–/– BMDMs (Fig. 5D). Overall, these data reveal that cell death pathways are major targets of the ubiquitination machinery during F. novicida infection and that IFN-I signaling is essential for the regulation of ubiquitination modifications.
Comments (0)