
記住我
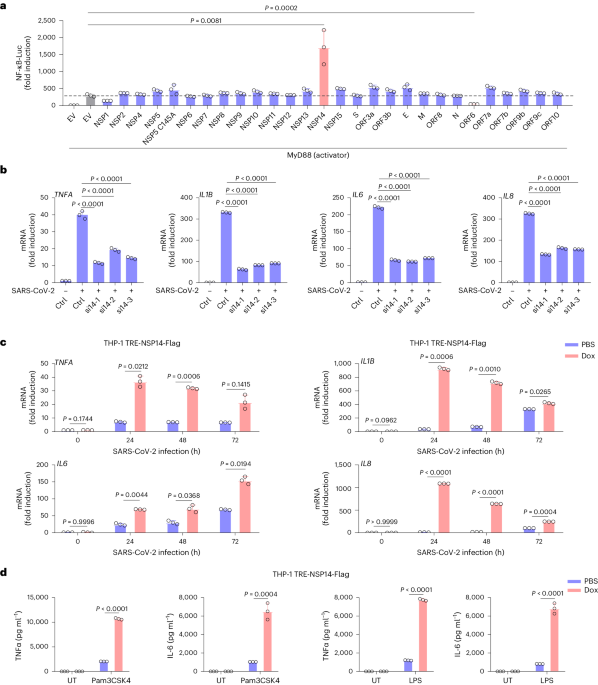
Peripheral nerves require neurotrophic factors for their maintenance38, but the source of such factors in the bone marrow is unknown. Analysis of published microarray data16 (National Center for Biotechnology Information (NCBI) accession number GSE33158) suggested that nerve growth factor (Ngf) was the only neurotrophic factor detected in adult bone marrow (Fig. 1a). Ngf expression was detected in Scf–GFP+CD45−Ter119−CD31− stromal cells, nearly all of which are LepR+(ref. 33), but little or no Ngf was detected in osteoblasts, endothelial cells or unfractionated whole bone marrow (WBM) cells (Fig. 1a). Similar results were obtained by RNA sequencing22 (NCBI accession number PRJNA914703), which detected Ngf in PDGFRα+CD45−Ter119−CD31− stromal cells, nearly all of which are LepR+(ref. 33), but not in endothelial cells or WBM cells (Fig. 1b).
Fig. 1: Ngf is mainly expressed in the bone marrow by LepR+ stromal cells.
a,b, The expression of neurotrophic factors by microarray analysis16 (a) and RNA sequencing22 (b) in bone marrow stromal cells (isolated on the basis of expression of Scf–GFP (a) or PDGFRα (b) staining, both of which are nearly completely overlapping with LepR expression33), VE-cadherin+ bone marrow endothelial cells, Col2.3–GFP+CD45−Ter119−CD31− osteoblasts, and WBM cells (three mice in a and two mice in b, from three or two independent experiments, respectively). c, Uniform manifold approximation and projection (UMAP) plot showing clustering of single-cell RNA sequencing analysis of 4,209 non-haematopoietic cells from enzymatically dissociated bones/bone marrow in 8-week-old mice39. d, Ngf is mainly expressed by Lepr+ stromal cells (cell cluster 11 in c) and smooth muscle cells (cell cluster 12). e, Ngf expression by all cell clusters shown in c (cells were obtained from four mice and analysed in three independent experiments). f, Ngf expression by qRT–PCR in LepR+CD45−Ter119−CD31− stromal cells, NG2–DsRed+ smooth muscle cells, Col1a1–GFP+ osteoblasts, VE-cadherin+ endothelial cells and unfractionated cells from the bone marrow of 2-month-old mice (three mice from three independent experiments). g, Deep imaging of femur bone marrow from adult NgfmScarlet/+ mouse: the Ngf–mScarlet+ cells were found around endomucinhigh sinusoids (arrowhead) as well as around endomucinlow arterioles (arrow; the images are representative of five mice). h,i, Flow cytometric analysis of enzymatically dissociated bone marrow from NgfmScarlet/+ mice: 89% of Ngf–mScarlet+ cells were LepR+, and most LepR+ cells were Ngf–mScarlet+ (four mice from four independent experiments). SSC-A, side scatter area. All data represent mean ± standard deviation.
Single-cell RNA sequencing of enzymatically dissociated cells from the femurs and tibias of 8-week-old mice showed that most Ngf-expressing cells in adult bone marrow were LepR+ cells (Fig. 1c,d; NCBI accession number PRJNA835050)39. Ngf was also expressed by a much smaller number of SMA+NG2+ smooth muscle cells, and by rare osteoblasts (OLC-2 cells), Schwann cells and fibroblasts (Fig. 1c–e). Little or no Ngf was detected in endothelial cells, chondrocytes or other stromal cells (Fig. 1c–e). Quantitative reverse-transcription polymerase chain reaction (qRT-PCR) confirmed that Ngf was highly expressed by LepR+CD45−Ter119−CD31− stromal cells and SMA+NG2+ smooth muscle cells, with approximately 100-fold lower expression by Col2.3–GFP+CD45−Ter119−CD31− osteoblasts and no expression by endothelial cells or WBM cells (Fig. 1f). LepR+ cells and smooth muscle cells were thus the main sources of NGF in the bone marrow.
To identify the location of Ngf-expressing cells in adult bone marrow, we generated an Ngf–mScarlet (NgfmScarlet) knock-in reporter allele (Extended Data Fig. 1a–c). Confocal imaging40 of cleared femurs from adult NgfmScarlet/+ mice showed that Ngf–mScarlet was expressed by stromal cells surrounding Endomucinlow arterioles as well as Endomucinhigh sinusoids (Fig. 1g). While the peri-arteriolar staining appeared more prominent, the abundance of sinusoids throughout the bone marrow meant that most of the Ngf–mScarlet staining was peri-sinusoidal. The peri-arteriolar staining probably reflected Ngf–mScarlet expression by both peri-arteriolar LepR+Osteolectin+ cells36 as well as SMA+NG2+ smooth muscle cells (Fig. 1e).
Flow cytometric analysis of enzymatically dissociated bone marrow cells showed that 0.087 ± 0.029% of bone marrow cells were Ngf–mScarlet+ (Fig. 1h). Consistent with the single-cell RNA sequencing (Fig. 1d), 89 ± 5.3% of bone marrow Ngf–mScarlet+ cells were LepR+ (Fig. 1h) and 82 ± 10% of all bone marrow LepR+ cells were Ngf–mScarlet+ (Fig. 1i). The remaining ~10% of Ngf–mScarlet+ cells that were negative for LepR within the bone marrow were mainly SMA+NG2+ smooth muscle cells (Extended Data Fig. 1d,f). There were also rare osteoblasts (Extended Data Fig. 1e,g), Schwann cells (Extended Data Fig. 1h) and macrophages (Extended Data Fig. 1i,j) that were Ngf–mScarlet+. The flow cytometry gates used to sort each cell population characterized in this study are shown in Extended Data Fig. 2.
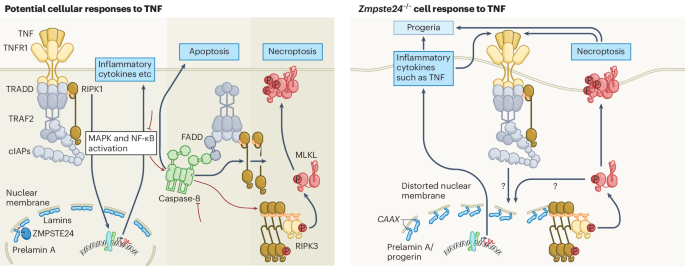
NGF from LepR+ cells is required for bone marrow innervationTo test if NGF is required for bone marrow innervation we generated mice with a floxed Ngf allele (Extended Data Fig. 3a–c). We conditionally deleted Ngf in LepR+ cells using Leprcre, in smooth muscle cells using NG2–creER, in osteoblasts using Col1a1–creER, and in Schwann cells using GFAP–cre. Deletion from smooth muscle cells, osteoblasts or Schwann cells had no significant effect on bone marrow NGF levels (Fig. 2a) or the number of nerve fibres in adult bone marrow (Fig. 2b and Extended Data Fig. 3d–g). Therefore, smooth muscle cells, osteoblasts and Schwann cells were not significant sources of NGF for nerve maintenance in bone marrow.
Fig. 2: NGF from LepR+ cells is necessary to maintain nerve fibres in the bone marrow.
a,b, NGF protein levels in bone marrow serum (a) and the area occupied by peripheral nerves in bone marrow (b) from 6–8-month-old Ngffl/∆ control (n = 8), Leprcre/+; Ngffl/∆ (n = 8), NG2–creER; Ngffl/∆ (n = 4), Col1a1–creER; Ngffl/∆ (n = 4), and GFAP–cre; Ngffl/∆ mice (n = 4) (four to eight mice per genotype from four independent experiments). c, Deep imaging of femur bone marrow from 6–8-month-old Leprcre/+; Ngffl/∆ and Ngffl/∆ littermate control mice (images are representative of three experiments with one mouse per genotype per experiment). d,e, Peripheral nerves were present in normal numbers in the bone marrow of 2-month-old Leprcre/+; Ngffl/∆ mice but were absent from the bone marrow of 6-month-old Leprcre/+; Ngffl/∆ mice (n = 6) (d) when the efficiency of Ngf deletion was more than 90% (n = 5) (e) (five to six mice per genotype per age from five to six independent experiments). f,g, Bone marrow and spleen cellularity (f) and haematopoietic stem and progenitor cell frequencies in the bone marrow (g) of 6–8-month-old Leprcre/+; Ngffl/∆ and Ngffl/∆ littermate control mice (six mice per genotype from six independent experiments). h,i, Defective circadian regulation of haematopoietic stem/progenitor cell mobilization into the blood of Leprcre/+; Ngffl/∆ mice based on numbers of LSK cells (h) and colony-forming progenitors (i) per millilitre of blood at different Zeitgeber times (ZT5, late morning; ZT13, just after nightfall; five mice per genotype from five independent experiments). All data represent mean ± standard deviation. The statistical significance of differences among treatments was assessed using a one-way ANOVA (a) followed by the Dunnett’s multiple comparisons adjustment, a Kruskal–Wallis test (b) followed by the Dunn’s multiple comparisons adjustment, Mann–Whitney tests (d and e) or Student’s t-tests (f and g) followed by the Holm–Šidák’s multiple comparisons adjustment, or matched samples two-way ANOVAs (h and i) followed by the Šidák’s multiple comparisons adjustment. All the statistical tests were two-sided. Not significant (NS): P > 0.05.
Adult Leprcre/+; Ngffl/∆ mice were born in expected numbers and did not differ from littermate controls in terms of gross appearance (Extended Data Fig. 4a), body length (Extended Data Fig. 4b) or body mass (Extended Data Fig. 4c). LepR+ cells from Leprcre/+; Ngffl/∆ mice had Ngf transcript levels that were approximately 30% of control levels at 2 months of age and less than 10% of control levels at 6 months of age (Fig. 2e). Deletion of Ngf from LepR+ cells profoundly depleted NGF from the bone marrow by 6 months of age (Fig. 2a).
No bone marrow innervation defect was apparent in Leprcre/+; Ngffl/∆ mice during development as the number of nerve fibres in the bone marrow was normal in 2-month-old Leprcre/+; Ngffl/∆ mice (Fig. 2d). However, by 6 months of age, when recombination in LepR+ cells was nearly complete, we observed virtually no nerve fibres in the bone marrow of Leprcre/+; Ngffl/∆ mice (Fig. 2b–d). Peripheral nerves appeared to be present in normal numbers in the quadriceps of 6-month-old Leprcre/+; Ngffl/∆ mice (Extended Data Fig. 4d–f). Nerve fibres thus grew into the bone marrow normally during development in Leprcre/+; Ngffl/∆ mice but became depleted within the bone marrow, but not outside of the bone marrow, by 6 months of age, when NGF was depleted to less than 10% of control levels in the bone marrow.
Consistent with prior studies4,6,11, loss of nerve fibres from the bone marrow did not have any gross effect on steady-state haematopoiesis. Six-month-old Leprcre/+; Ngffl/∆ mice did not differ from littermate controls in terms of bone marrow or spleen cellularity (Fig. 2f), or the frequencies of HSCs, multipotent haematopoietic progenitors (MPPs), granulocyte–macrophage progenitors (GMPs), megakaryocyte–erythroid progenitors (MEPs), common myeloid progenitors (CMPs) or common lymphoid progenitors (CLPs) in the bone marrow (Fig. 2g). There were also no differences in blood cell counts (Extended Data Fig. 4g–i) or in the frequencies of B220+ B cells, CD3+ T cells, Gr1+Mac1+ myeloid cells, CD41+ megakaryocyte lineage cells or CD71+/Ter119+ erythroid lineage cells in the bone marrow or spleen (Extended Data Fig. 4j–n). Finally, WBM cells from 6-month-old Leprcre/+; Ngffl/∆ mice and littermate controls did not differ in their capacity to reconstitute myeloid, B or T cells upon competitive transplantation into irradiated mice (Extended Data Fig. 4o–r). Bone marrow nerve fibres thus appear to be dispensable for normal adult haematopoiesis.
In agreement with earlier studies6,11, we did observe a defect in the circadian mobilization of Lineage−Sca1+c-kit+ (LSK) haematopoietic stem/progenitor cells (Fig. 2h) and colony-forming progenitors (Fig. 2i) into the blood during midmorning (Zeitgeber Time 5) in 6-month-old Leprcre/+; Ngffl/∆ as compared with littermate control mice.
Bone marrow innervation promotes haematopoietic regenerationTo test for haematopoietic regeneration defects, we lethally irradiated (1,080 rads) and transplanted a radioprotective dose of 1,000,000 WBM cells into 6-month-old LeprCre/+; Ngffl/∆ mice and littermate controls. While all control mice survived, 41% (9 of 22) of LeprCre/+; Ngffl/∆ mice died between 10 and 18 days after irradiation, consistent with haematopoietic failure (Fig. 3a). The LeprCre/+; Ngffl/∆ mice exhibited significantly lower white blood cell (WBC, Fig. 3b), red blood cell (RBC, Fig. 3c) and platelet (PLT) counts (Fig. 3d) as well as bone marrow cellularity (Fig. 3e) and LSK cell numbers (Fig. 3f) at 14 and 28 days after irradiation. At 28 days after irradiation, HSC numbers were much lower in the bone marrow of LeprCre/+; Ngffl/∆ as compared with littermate control mice (Fig. 3g). We thus observed broad reductions in blood and bone marrow cell counts as well as the numbers of haematopoietic stem and progenitor cells in LeprCre/+; Ngffl/∆ mice at 14–28 days after irradiation.
Fig. 3: Leprcre/+; Ngffl/∆ mice exhibit defects in haematopoietic and vascular regeneration after irradiation.
a, Survival of Leprcre/+; Ngffl/∆ and Ngffl/∆ littermate control mice after irradiation and transplantation of wild-type bone marrow cells (22 mice per genotype from 3 independent experiments). b–d, WBC (b), RBC (c) and PLT (d) counts from Leprcre/+; Ngffl/∆ and littermate control mice before (D0) and 7, 14 and 28 days after irradiation. e,f, Bone marrow cellularity (e) and LSK cell numbers (f) from 6-month-old Leprcre/+; Ngffl/∆ and littermate control mice on day (D)0, D7, D14 and D28 after irradiation. g, Numbers of HSCs in bone marrow (always one tibia and one femur) from Leprcre/+; Ngffl/∆ and littermate control mice before and 28 days after irradiation. h–k, Cellularity (h) and numbers of LSK cells, (i) LepR+ cells (j) and endothelial cells (k) in the bone marrow of Leprcre/+; Ngffl/∆ and littermate control mice on D10 after irradiation (a total of 12 mice per genotype from 3 independent experiments). l,m, Leakage of intravenously injected Evans blue dye into femur bone marrow at D10 (l), D14 and D28 (m) after irradiation of Leprcre/+; Ngffl/∆ and littermate control mice. n, Endomucin staining of the vasculature in the bone marrow of Leprcre/+; Ngffl/∆ and littermate control mice 28 days after irradiation (representative of three experiments). o,p, The percentages of LepR+ cells (o) and endothelial cells (p) from Leprcre/+; Ngffl/∆ and littermate control mice that incorporated a 48-h pulse of bromodeoxyuridine (BrdU) 28 days after irradiation. q,r, Numbers of LepR+ cells (q) and endothelial cells (r) in the bone marrow 28 days after irradiation. s, Survival of 6-month-old Leprcre/+; Ngffl/∆ and Ngffl/∆ littermate control mice after 5-FU treatment (19 Leprcre/+; Ngffl/∆ mice and 15 Ngffl/∆ mice in 3 independent experiments). t–v, Cellularity (t), numbers of HSCs (u) and LSK cells (v) in bone marrow from 6-month-old Leprcre/+; Ngffl/∆ and littermate control mice 12 days after 5-FU treatment. Unless otherwise specified, each panel shows five mice per genotype from five independent experiments per timepoint. All data represent mean ± standard deviation. Statistical significance was assessed using log-rank tests (a and s), two-way ANOVAs (m and p) or matched samples two-way ANOVAs (b and d) followed by the Šidák’s multiple comparisons adjustment, Student’s t-tests (h, l and t) or Student’s t-tests (c, e–g, o, q, r, u and v) or Welch’s t-tests (i–k) followed by the Holm–Šidák’s multiple comparisons adjustment. All statistical tests were two-sided. Not significant (NS) means P > 0.05.
We also assessed vascular and stromal cell regeneration in LeprCre/+; Ngffl/∆ and littermate control mice. At 10 days after lethal irradiation and transplantation, we observed significantly reduced numbers of bone marrow cells (Fig. 3h), LSK cells (Fig. 3i), LepR+ stromal cells (Fig. 3j) and endothelial cells (Fig. 3k), as well as increased vascular leakage (Fig. 3l) in the bone marrow of LeprCre/+; Ngffl/∆ mice as compared with littermate controls. At 28 days after irradiation, blood vessels were patent in control mice but remained leaky (Fig. 3m) and morphologically abnormal (Fig. 3n) in LeprCre/+; Ngffl/∆ mice. LeprCre/+; Ngffl/∆ mice had significantly less proliferation by LepR+ cells at 14 days after irradiation (Fig. 3o) and by endothelial cells at 14 and 28 days after irradiation (Fig. 3p). In this experiment we observed trends towards reduced numbers of LepR+ cells and endothelial cells at 14 days after irradiation and significant reductions in the numbers of these stromal cells at 28 days after irradiation (Fig. 3q,r). The loss of nerve fibres from the bone marrow in LeprCre/+; Ngffl/∆ mice was thus associated with broad defects in the regeneration of haematopoietic, stromal and vascular cells at 10–28 days after irradiation.
To test if defects in haematopoietic regeneration were also evident after sublethal irradiation, we administered 650 rads to 6-month-old LeprCre/+; Ngffl/∆ and littermate control mice. The LeprCre/+; Ngffl/∆ mice exhibited significantly reduced survival from 12 to 15 days after irradiation (Extended Data Fig. 5a) as well as reduced bone marrow cellularity (Extended Data Fig. 5b), HSC numbers (Extended Data Fig. 5c) and LSK cell numbers (Extended Data Fig. 5d) as compared with littermate controls at 28 days after irradiation.
To test if defects in haematopoietic, vascular and stromal cell regeneration were evident after chemotherapy, we treated LeprCre/+; Ngffl/∆ and littermate control mice with 5-fluorouracil (5-FU). The LeprCre/+; Ngffl/∆ mice exhibited significantly reduced survival from 12 to 15 days after irradiation (Fig. 3s) as well as reduced numbers of bone marrow cells (Fig. 3t), HSCs (Fig. 3u) and LSK cells (Fig. 3v) as compared with littermate controls at 12 days after 5-FU treatment. Impaired haematopoietic regeneration was thus observed in LeprCre/+; Ngffl/∆ mice irrespective of whether myeloablation was induced by 5-FU treatment, sublethal irradiation or lethal radiation.
NGF acts locally to promote nerve maintenance in bone marrowNeurotrophic factors act locally to promote the survival of innervating neurons41,42. To test if NGF acts locally within the bone marrow to promote nerve fibre maintenance, we deleted Ngf using Prx1–cre. Prx1–cre recombines in limb mesenchymal cells, including in LepR+ cells that form in the bone marrow of limb bones, but not within the axial skeleton33,39,43. Two-month-old Prx1–cre; Ngffl/fl mice exhibited a lack of nerve fibres in femur bone marrow but normal innervation of vertebral bone marrow (Fig. 4a–c). This demonstrates that NGF acts locally within the bone marrow to promote nerve fibre maintenance.
Fig. 4: Prx1–cre; Ngffl/fl mice exhibit a loss of nerve fibres as well as defects in haematopoietic and vascular regeneration in long bones but not in vertebrae.
a,b, Nerve fibres (green) were visible in femur (a) and vertebra (b) bone marrow from Ngffl/fl control mice and in vertebra bone marrow (b) from 2-month-old Prx1–cre; Ngffl/fl mice but not in femur bone marrow (a) from Prx1–cre; Ngffl/fl mice (representative of three independent experiments). c, The area occupied by peripherin+ nerve fibres in bone marrow sections from 2-month-old Prx1–cre; Ngffl/fl and littermate control mice. d–f, Under steady-state conditions, 2-month-old Prx1–cre; Ngffl/fl mice did not significantly differ from littermate control mice in terms of spleen, femur bone marrow or vertebral bone marrow cellularity (d), the frequencies of haematopoietic stem and progenitor cell populations in femur bone marrow (e) (six mice per genotype from six independent experiments in c–e), or the levels of donor cell reconstitution upon competitive transplantation into irradiated mice (f, femur bone marrow cells from five donor mice were transplanted into a total of five recipients per donor per genotype in five independent experiments). g–k, At 28 days after irradiation, Prx1–cre; Ngffl/fl and littermate control mice did not significantly differ in terms of bone marrow cellularity (g) and the numbers of HSCs (h), LSK cells (i), LepR+ cells (j) or endothelial cells (k) in the vertebrae, but all of these parameters were significantly lower in femur bone marrow (six mice from six independent experiments). l, Leakage of intravenously injected Evans blue dye into femur and vertebra bone marrow 28 days after irradiation (five mice from five independent experiments). All data represent mean ± standard deviation. The statistical significance of differences among treatments was assessed using Mann–Whitney tests (c), Student’s t-tests (d, e and l) or Welch’s t-tests (i) followed by the Holm–Šidák’s multiple comparisons adjustment, or matched samples two-way ANOVAs (f–h, j and k) followed by the Šidák’s multiple comparisons adjustment. All the statistical tests were two-sided. Not significant (NS): P > 0.05.
Consistent with the phenotype observed in LeprCre/+; Ngffl/∆ mice, Prx1–cre; Ngffl/fl mice exhibited normal bone marrow haematopoiesis under steady-state conditions but impaired haematopoietic and vascular regeneration in limb bones. Compared with littermate controls, Prx1–cre; Ngffl/fl mice exhibited normal femur bone marrow, vertebral bone marrow and spleen cellularity (Fig. 4d), and normal frequencies of HSCs and restricted haematopoietic progenitors (Fig. 4e) in femur bone marrow. WBM cells from the femurs of Prx1–cre; Ngffl/fl mice and littermate controls did not differ in their capacity to reconstitute myeloid, B or T cells upon competitive transplantation into irradiated mice (Fig. 4f and Extended Data Fig. 5e–g).
To assess the regeneration of haematopoiesis after irradiation, we lethally irradiated (1,080 rads) and transplanted a radioprotective dose of 1,000,000 WBM cells into 2-month-old Prx1–cre; Ngffl/fl mice and littermate controls. At 28 days after irradiation, the regeneration of bone marrow cellularity (Fig. 4g), HSCs (Fig. 4h), LSK cells (Fig. 4i), LepR+ cells (Fig. 4j) and endothelial cells (Fig. 4k) were all significantly impaired in femur, but not vertebral, bone marrow in Prx1–cre; Ngffl/fl mice. Consistent with this, femur, but not vertebral, bone marrow blood vessels in Prx1–cre; Ngffl/fl mice were leaky at 28 days after irradiation (Fig. 4l). Blood cell counts did not significantly differ between Prx1–cre; Ngffl/fl and littermate control mice before or after irradiation (Extended Data Fig. 5h–j), consistent with the observation that haematopoietic regeneration was impaired only in limb bones. NGF thus acts locally within the bone marrow to promote haematopoietic, stromal and vascular cell regeneration.
Nerve sprouting increases regeneration factor expressionWhen compared with non-irradiated bone marrow, we observed a significant increase in NGF levels at 14 days after irradiation in 6-month-old control mice but not in 6-month-old LeprCre/+; Ngffl/∆ mice (Fig. 5a). In the bone marrow, Ngf–mScarlet was mainly expressed by adipocytes (Fig. 5b) and LepR+ cells (Fig. 5c and Extended Data Fig. 6a) at 14 days after irradiation. Little or no Ngf–mScarlet expression was observed among haematopoietic/endothelial cells or LepR negative stromal cells in the bone marrow (Fig. 5c). By qRT–PCR, Ngf levels were similar in adipocytes and in LepR+ cells (Fig. 5d).
Fig. 5: NGF from LepR+ cells and adipocytes promotes nerve sprouting after irradiation, increasing the expression of regeneration factors.
a, NGF in bone marrow serum from 6–8-month-old Leprcre/+; Ngffl/∆ and Ngffl/∆ littermate controls before (n = 3 mice per genotype), or 14 (n = 4) or 28 (n = 4) days after irradiation and transplantation of radioprotective wild-type bone marrow cells (three to four independent experiments per timepoint). D, day. b, Perilipin+ adipocytes in a 30-µm-thick section from NgfmScarlet/+ femur bone marrow were positive for Ngf–mScarlet (representative of three experiments). There are also Ngf-expressing LepR+ cells in this image (Extended Data Fig. 6a). c, Flow cytometric analysis of enzymatically dissociated bone marrow from NgfmScarlet/+ mice 14 days after irradiation (four mice from four independent experiments). d, Ngf expression by qRT–PCR (three mice (WBM or LepR+ cells) or five mice (adipocytes) from three independent experiments). e, The area occupied by peripherin+ nerve fibres in bone marrow sections from 6–8-month-old Leprcre/+; Ngffl/∆ and littermate control mice (five mice per genotype per timepoint from five independent experiments). f,g, Representative images (f) and quantification (g) showing that nerve fibres (red) in femur bone marrow sections from 6–8-month-old Wnt1–Cre; Rosa26tdTomato mice before or at 14 or 28 days after irradiation (five mice per timepoint from five independent experiments). h–j, SCF (h), VEGF (i) and Ang2 (j) in bone marrow serum from 6–8-month-old Leprcre/+; Ngffl/∆ and littermate control mice (eight mice per genotype per timepoint from eight independent experiments). k–n, The β2 agonist salbutamol (Salb.) rescued the regeneration of bone marrow cellularity (n = 6) (k) and the numbers of HSCs (n = 6 mice per treatment) (l) and LSK cells (n = 6) (m) as well as the patency of the vasculature (n = 5) (n) in 6–8-month-old Leprcre/+; Ngffl/∆ mice at 28 days after irradiation (five to six independent experiments). o, Western blot of protein from LepR+ cells isolated from Leprcre/+; Ngffl/∆ and littermate control mice at 14 days after irradiation and transplantation (representative of three independent experiments). All data represent mean ± standard deviation. Statistical significance was assessed using two-way ANOVAs followed by Tukey’s (a and h) or Šidák’s (k–n) multiple comparisons adjustments, Mann–Whitney (e) or Student’s t-tests (i and j) followed by Holm–Šidák’s multiple comparisons adjustments for comparisons between mutants and controls, or one-way ANOVAs (e, g, i and j) followed by Šidák’s multiple comparisons adjustments for comparisons between timepoints. All statistical tests were two-sided. Not significant (NS): P > 0.05.
We observed significantly increased nerve fibre density in control bone marrow after irradiation but not in the bone marrow of 6-month-old LeprCre/+; Ngffl/∆ mice (Fig. 5e). We irradiated Wnt1–cre; Rosa26–tdTomato mice, which express Tomato in neural crest-derived cells, including nerve fibres and Schwann cells44. Nerve fibres were much more abundant in the bone marrow at 14 days after irradiation as compared with non-irradiated controls (Fig. 5f,g). Nerve fibres were closely associated with arterioles under steady-state conditions and after irradiation (Extended Data Fig. 6b). By 28 days after irradiation, when NGF levels in the bone marrow returned nearly to normal (Fig. 5a), the density of nerve fibres also returned nearly to normal (Fig. 5g).
To test if the increase in NGF levels in the bone marrow after irradiation caused nerve fibre sprouting, we examined 4–5-month-old LeprCre/+; Ngffl/∆ mice. These mice had lower levels of NGF in the bone marrow as compared with littermate controls (Extended Data Fig. 6c). They had a normal density of nerve fibres in the bone marrow before irradiation but, unlike control mice, did not exhibit an increase in nerve fibres 14 days after irradiation (Extended Data Fig. 6d). These 4–5-month-old LeprCre/+; Ngffl/∆ mice exhibited delayed regeneration of bone marrow cellularity (Extended Data Fig. 6e), HSCs (Extended Data Fig. 6f) and LSK cells (Extended Data Fig. 6g) as compared with littermate controls. Therefore, the sprouting of nerve fibres in the bone marrow after irradiation occurs in response to increased NGF production by LepR+ cells, and the adipocytes they give rise to, and this accelerates haematopoietic regeneration.
We hypothesized that bone marrow nerve fibres increased the production of growth factors that promote haematopoietic and vascular regeneration. We found by enzyme-linked immunosorbent assay analysis that SCF (Fig. 5h), VEGF (Fig. 5i) and Ang2 (Fig.
留言 (0)