
Remember me




In this study, we recruited a total of 158 patients who had been diagnosed with LUAD and divided them into two cohorts. The clinical data for both cohorts are presented in Table 1. Cohort 1 was designated as the testing cohort and included 30 patients with early-stage LUAD confirmed through pathological examination. Each patient provided a lung cancer tissue sample along with an adjacent non-cancerous tissue sample, resulting in a collection of 60 resected specimens that underwent high-throughput transcriptome sequencing. The obtained sequencing data were comprehensively analyzed together with the corresponding pathological and imaging information of the patients. Cohort 2, known as the validation cohort, consisted of 128 LUAD patients from whom matched pathological tissue samples and partial follow-up data were collected for laboratory testing and survival analysis purposes.
Table 1 Clinical information of included patientsThe study design of our research was presented in Supplementary Figure S1, which illustrates the experimental framework. The analysis of differentially expressed genes (DEGs) was conducted on the transcriptome sequencing data obtained from patients in Cohort 1. A total of 2161 DEGs were identified, consisting of 1827 known genes and a subset of hypothetical genes (Fig. 1a and b). These genes exhibited distinct expression patterns during the transition from normal tissue to tumor tissue at early stages of LUAD onset, thus referred to as oncogenesis-related DEGs (ORDEGs). To provide a comprehensive understanding of the enriched functions represented by these ORDEGs, we employed WGCNA to establish a hierarchical clustering tree for them and utilized the dynamic-cut method to merge them into ten gene-network modules.In the co-expression network heatmap, genes within each module showed a high level of expression similarity, whereas those belonging to different modules demonstrated low similarity (Fig. 1c). Following that, correlation analysis was performed to clarify the relationships among various gene modules (Supplementary Fig. S2a and b). The results indicated a significant positive correlation between the brown and green modules (r = 0.73, P = 0.01). Additionally, there was an evident correlation observed for genes present in both brown and green modules concerning the turquoise module. These results indicated that WGCNA effectively clustered genes with high similarity into the same gene module, suggesting the establishment of a co-expression network among genes. Furthermore, there was a strong correlation between different gene modules. Subsequently, we utilized the Metascape Database to conduct functional enrichment analysis of DEGs and explored their functional interplay across various modules. We performed comprehensive functional enrichment analysis of all DEGs and identified gene subsets with similar GO terms (Fig. 1d). As expected, the significant enrichments were observed in cell cycle regulation, cell proliferation, and related pathways such as the mitogen-activated protein kinase pathway. To gain a more comprehensive understanding of the functional correlations within each gene module, we conducted functional enrichment analysis across the various gene-network modules. Our findings revealed significant enrichment for top-level GO biological processes, such as metabolic process (Fig. 1e), in the turquoise, blue, brown, and green gene modules. Subsequent correlation analysis between gene significance and module membership values revealed a positive correlation between the genes within each respective module and their corresponding functional characteristics (Supplementary Fig. S2c-f), indicating that the enriched genes in each module can effectively represent its specific functions. Furthermore, we conducted functional enrichment analysis of DEGs within their respective modules using the Metascape Database (Fig. 1f-i). It was noteworthy that specific DEGs were observed to be enriched in plasma functions, such as the apical plasma membrane within the turquoise module (Fig. 1f), anchored components of the membrane within the brown module (Fig. 1h), and apicolateral plasma membrane within the green module (Fig. 1i). Considering these findings, it could be inferred that the reorganization of cellular membrane constituents an integral part of metabolic processes and was synchronized with rapid cell proliferation to facilitate oncogenesis progression from lung preneoplasia to adenocarcinoma.
Fig. 1
WGCNA and functional annotations for DEGs in different modules. a The heatmap showed the DEGs between 30 paracancerous tissues and 30 tumour tissues. b The volcano plot showed the DEGs between 30 paracancerous tissues and 30 tumour tissues. The red dots represented up-regulated significant DEGs and the blue dots represented sown-regulated significant DEGs. c WGCNA network heatmap showed cluster dendrogram and genetic similarity among different modules. d Functional annotations for 1827 ORDEGs. e Top-level gene ontology biological processes enrichment of turquoise module、blue module、brown module and green module. f Functional annotations for ORDEGs in turquoise module. g Functional annotations for ORDEGs in blue module. h Functional annotations for ORDEGs in brown module. i Functional annotations for ORDEGs in green module
Enhancement of cell cycle promoted the transition of early-stage LUAD from non-invasiveness to invasivenessIt well known that pathological diagnosis and radiological imaging signs are important clinical index for early-stage LUAD diagnosis. To further understand the relationships between the molecular characteristics of early-stage LUAD and their different clinical phenotypes, we collected postoperative pathological diagnoses and preoperative imaging data from the 30 patients in Cohort 1 (Fig. 2a, Supplementary Table S1). Among these patients, two were diagnosed with severe atypical adenomatous hyperplasia (AAH), three with adenocarcinoma in situ (AIS), three with minimally invasive adenocarcinoma (MIA), while the remaining twenty-two patients were diagnosed with invasive adenocarcinoma (IAC). Given that AAH, AIS, and MIA lesions exhibit non-invasiveness or low invasiveness, the eight patients demonstrating these clinicopathologic characteristics were classified as the non-invasive group. The remaining 22 patients with IAC lesions were categorized as the invasive group. Subsequently, a statistical analysis was conducted to examine the correlations between imaging features and pathological diagnosis in a cohort of 30 early-stage LUAD cases. Imaging characterization of invasive nodules revealed larger tumor diameters, higher consolidation tumor (C/T) ratios, and increased indications of vascular convergence and pleural indentation when compared to their non-invasive counterparts (Fig. 2b-f). These findings demonstrated a consistent correlation between the pathological characteristics and imaging features of invasive lesions. Initially, gene expression profiles were analyzed to compare the non-invasive group with the invasive group, resulting in 548 differentially expressed genes (DEGs) that were highlighted. By intersecting these DEGs with ORDEGs, we identified 256 DEGs, out of which 169 were jointly up-regulated and 87 were jointly down-regulated. These DEGs were closely associated with the invasiveness of early-stage LUAD (Fig. 2g-i). Functional analysis of these 256 DEGs revealed a significant enrichment in mitosis and cell-cycle processes (Fig. 2j). A pathway diagram illustrating the cell cycle was constructed using the KEGG Database, revealing that a majority of differentially expressed genes (DEGs) were predominantly enriched in the S and G2/M phases of the cell cycle. These DEGs included essential components involved in replication forks during the S phase, such as ORC1, ORC6, CDC6, MCM2, MCM4, and MCM6. Furthermore, the components associated with spindle assembly checkpoint regulation during the M phase were also identified among these DEGs, including BUB1B, BUB1, PLK1, and PKMYT (Fig. 2k). Additionally, the marker genes representing distinct phases of G1, S, G2, and M were selected from the Molecular Signatures Database for both experimental groups and visualized on a heatmap (Fig. 2l). Next, single-sample gene set enrichment analysis (ssGSEA) was conducted on the gene sets according to previously described methods [27]. The results demonstrated a significant elevation in ssGSEA score within the invasive group compared to the non-invasive group (Fig. 2m). In conclusion, our findings suggested that augmented cell cycle activity might facilitate the transition from a non-invasive to an invasive state in early-stage LUAD.
Fig. 2
Cell-cycle activation was the primary feature of early-stage LUAD onset and progression. a Representative image showed the pathological and radiological features of patients in cohort 1. b-e Statistic analyses of tumour diameters, C/T ratios, vascular convergence sign and pleural indentation sign in patients with non-invasive and invasive nodes. *P < 0.05, **P < 0.01, ****P < 0.0001, the student’s t test. f Pie chart showed the percentage of patients with different pathological and radiological features. g Venn showed 169 common up regulated DEGs between ORDEGs and DEGs in non-invasive versus invasive group. h Venn showed 87 common down regulated DEGs between ORDEGs and DEGs in non-invasive versus invasive group. i The heatmap showed the DEGs between non-invasive and invasive tissues. j Functional annotations for 256 common DEGs in Fig. 3G-H. k Enrichment analysis of cell cycle pathway in KEGG database. l The heatmap showed the marker genes in G1, S, G2, M phase between non-invasive and invasive tissues. m Statistic analysis of ssGSEA z-scores in G1, S, G2, M phase marker genes between non-invasive and invasive tissues. *P < 0.05, ns: no significance, the student’s t test
Lipid rafts-driven invasiveness of early-stage LUAD was relative to the activation of cell-cycleTo gain novel insights into the underlying relationship connecting different gene signatures related to the cell cycle and alterations in the plasma membrane, a series of transcriptomic profile-based analytical methods were employed. As depicted in Fig. 1e, we observed an enrichment of metabolic processes during the initiation and progression of early-stage LUAD. Considering that metabolic processes were implicated in plasma membrane synthesis and cellular proliferation, our initial analysis focused on exploring the correlation between gene expression profiles and metabolic phenotypes stratified based on imaging characteristics in early-stage LUAD. To begin with, assessing vascular coverage through imaging served as an indicator for determining a tumor's capacity to obtain metabolites due to angiogenesis providing enough nutrients that supported its growth [28]. Out of a total of 22 IACs cases examined in this study, 6 displayed negative indications of vascular convergence while 16 demonstrated positive indications (Fig. 3a and b). Additionally, considering that standard uptake value (SUV) obtained from positron emission tomography-computed tomography (PET-CT) was also regarded as a clinical parameter used for evaluating tumor metabolism status; we gathered PET-CT data from 7 IACs cases. By analyzing both SUV values and indicators related to vascular convergence patterns among these cases; they were categorized into two distinct metabolic groups: 3 individuals fell under the low-metabolic group characterized by low SUV values along with negative indications of vascular convergence; conversely 4 individuals belonged to the high-metabolic group displaying elevated SUV values alongside positive indications of vascular convergence patterns (Fig. 3c and d). Then, a total of 475 differentially expressed genes (DEGs) were identified from the gene expression analysis (Fig. 3e). Subsequent GO enrichment analysis revealed characteristics closely associated with lipid rafts, such as membrane raft, membrane microdomain, signaling receptor regulator activity, and glycerophospholipid biosynthesis (Fig. 3f). Notably, significant alterations in mitosis were observed. These changes in the cell plasma membrane related to lipid raft formation aligned with the aforementioned findings observed during the progression from pre-invasive to minimally invasive and invasive LUAD. The available evidence suggests that inhibition of lipid raft formation may serve as a potential strategy to counteract tumor cell malignant proliferation [29]. To validate the relevance of lipid rafts and the cell cycle, we constructed a protein–protein interaction network of 475 DEGs using the molecular complex detection (MCODE) algorithm [30]. Ten densely connected network components were annotated (Fig. 3g). Among them, network component MCODE1, which was related to lipid raft-related signaling receptor regulator activity, showed significant interactions with cell cycle-related processes such as DNA glycosylase recognition binding (network component MCODE2) and anaphase-promoting complex (network component MCODE3). Interestingly, these interconnected network components align with the previously observed changes in the cell cycle (Fig. 2k). Furthermore, there was a significant positive correlation between genes related to lipid rafts and glycerophospholipid metabolism as well as genes associated with the cell cycle (Supplementary Fig. S3).
Fig. 3
Lipid rafts drove cell-cycle activation and invasiveness in early-stage LUAD. a Representative image of patients with or without vascular convergence sign. b Pie chart showed the percentage of patients with or without vascular convergence sign in invasive group. c Representative PET-CT images of patients in low SUV and high SUV group. d Statistic analysis of patients’ maximum SUV in low SUV and high SUV group. **P < 0.01, the student’s t test. e The heatmap showed the DEGs between low SUV tissues and high SUV tissues. f Functional annotations for 475 DEGs between low SUV tissues and high SUV tissues. g Protein–protein interaction network among 10 densely connected components calculated by MCODE algorithm. h Representative images of IHC assay detecting levels of caveolin-1 expression in paracancerous and tumour tissues of LUAD patients in cohort 2. i Statistic analysis of caveolin-1 AOD values in paired paracancerous and tumour tissues of stage I LUAD patients in cohort 2, n = 38. Bars, SD; ****P < 0.0001; the student’s t test. j Statistic analysis of caveolin-1 AOD values in paired paracancerous and tumour tissues of stage I A2 patients in cohort 2, n = 9. Bars, SD; **P < 0.01, the student’s t test. k Statistic analysis of caveolin-1 AOD values in paired paracancerous and tumour tissues of stage I A3 patients in cohort 2, n = 14. Bars, SD; ****P < 0.0001, the student’s t test. l Statistic analysis of caveolin-1 AOD values in paired paracancerous and tumour tissues of stage I B patients in cohort 2, n = 15. Bars, SD; ***P < 0.001, the student’s t test. m Correlation analysis of caveolin-1 AOD fold changes and Ki-67 positive percentages in paracancerous and tumour tissues of stage I LUAD patients, n = 38. **P < 0.01; spearman correlation test
Next, in order to further validate the involvement of lipid raft formation in LUAD, we collected tissue samples from 48 patients with LUAD and their corresponding para-carcinoma tissues. Among these samples, 38 pairs were obtained from patients diagnosed with stage I LUAD. The results obtained from the tissue microarray analysis revealed significantly higher levels of caveolin-1 expression, a well-established marker for lipid rafts, in early-stage LUAD tissues compared to para-carcinoma tissues (Fig. 3h and i). Moreover, when categorizing patients with stage I lung adenocarcinoma (n = 38), we observed that IB stage LUAD tissues (n = 15) exhibited the highest level of caveolin-1 expression as compared to IA2 (n = 9) and IA3 stage LUAD tissues (n = 14) (Fig. 3j-l). Additionally, our findings demonstrated a positive correlation between Ki-67 expression, which serves as a hallmark for cell proliferation, and caveolin-1 expression in early-stage LUAD tissues (r = 0.5, P < 0.01; Fig. 3m). These observations strongly suggested that enhanced lipid raft formation might drive the progression of early-stage LUAD through activation of the cell cycle machinery.
Integrative analysis of transcriptomics and lipidomics implicated the involvement of PLPP2 in lipid raft formation during early-stage LUADTo investigate the key determinant underlying the formation of lipid rafts in early-stage LUAD, we initially extracted 2408 DEGs from the TCGA dataset, comprising 78 patients with stage I LUAD. By intersecting these genes with 1827 ORDEGs from Cohort 1, an additional set of 114 lipid raft-related genes, and 77 glycerophospholipid metabolism-related genes from MSigDB, we successfully identified a singular common gene known as PLPP2 (Fig. 4a). Correlation analysis conducted on samples from Cohort 1 revealed that the expression of PLPP2 exhibited a concurrent increase alongside lipid raft-associated genes such as FLOT1, CLN3, SLC2A1, and HMOX (Fig. 4b and Supplementary Fig. S4). Furthermore, gene set enrichment analysis (GSEA) demonstrated significant enrichment of PLPP2 in membrane-associated functional subsets including organelle-inner membrane and protein insertion into the membrane (Fig. 4c and d). In order to ascertain whether PLPP2 conferred to the formation of lipid raft in LUAD cells, we conducted quantitative lipidomics analysis after overexpressing PLPP2 in NCI-H1299 cells. Our analysis identified a total of 1376 lipid species, including 213 differential metabolites. Among these, 86 metabolites were found to be up-regulated while 127 were down-regulated (Fig. 4e, Supplementary Table S2). Furthermore, KEGG analysis revealed significant enrichment of signaling pathways associated with lipid raft formation, such as cholesterol metabolism and glycerophospholipid metabolism (Fig. 4f). The classification of lipid components showed that the overexpression of PLPP2 in LUAD cells could significantly increase the metabolites associated with lipid raft formation, including cholesterol esters (CEs), sphingomyelin (SM), and sphingosine (SPH) (Fig. 4g). Additionally, quantitative analysis also revealed a significant increase in the contents of CE (18:0), SM (d18:0/22:0), SM (d18:1/17:1), SPH (d17:1), SPH (d18:1), and SPH (d18:2) in NCI-H1299 cells overexpressing PLPP2 (Fig. 4h). Overall, these findings suggested that PLPP2 might participate in altering the classification and content of lipid raft components such as CE, SM, and SPH to contribute to the formation of lipid rafts in LUAD cells.
Fig. 4
Transcriptomics and lipidomics analyses implicated PLPP2 in lipid raft formation during early-stage LUAD. a Venn showed common gene PLPP2 among 4 gene sets. Set 1: lipid raft gene set from molecular signatures database (MSigDB); set 2: glycerophospholipid metabolism gene set from MSigDB; set 3: ORDEGs; set 4: DEGs of paracancerous and tumour tissues of stage I LUAD patients in TCGA database. b Correlation analysis of PLPP2 and flotillin-1 levels in tumour tissues of patients in cohort 1, n = 30. **P < 0.01; spearman correlation test. c-d GSEA of PLPP2 levels in 30 tumour tissues of patients in cohort 1. e The heatmap showed differential metabolites between NCI-H1299 vector cells and PLPP2 OE cells. f KEGG annotation of differential metabolites between NCI-H1299 vector cells and PLPP2 OE cells. g Scatter plot showed the content differences of different subclasses of lipids in two groups of samples. Each point in the figure represented a type of lipid, and different colors represented different lipid subclasses. h Radar charts showed the levels of indicated lipids in NCI-H1299 vector cells and PLPP2 OE cells. *P < 0.05, **P < 0.01, ****P < 0.0001; the student’s t test
The elevated PLPP2 was an independent prognostic risk factor for early-stage LUAD patientsTo further confirm the aforementioned findings, we employed qRT-PCR to confirm the upregulation of PLPP2 in Cohort 1 samples. As shown in Fig. 5a, the expression levels of PLPP2 mRNA were significantly elevated in LUAD tissues compared to adjacent non-cancerous tissues. Subsequently, we assessed the expression levels of PLPP2 in a panel of LUAD cells using both qRT-PCR and western blotting. Compared to BEAS-2B cells, a normal human bronchial epithelial cell line, the levels of PLPP2 in LUAD cells were significantly increased (Fig. 5b and c). To evaluate the prognostic value of the PLPP2 for LUAD patients, we used the tissues microarray to detect the levels of PLPP2 in 80 patients who was available for follow-up information in Cohort 2 (Fig. 5d). Initially, we examined the expression of PLPP2 in 42 out of the total 80 cases with stage I LUAD and discovered that it was significantly higher compared to adjacent non-cancerous tissues (Fig. 5e and f). Additionally, consistent levels of PLPP2 were observed across different TNM stages in all LUAD patients, indicating its potential as a marker for malignancy (Supplementary Fig. S5a-b). To evaluate the predictive performance, we conducted a ROC analysis and calculated the area under the ROC curves. The results revealed that PLPP2 exhibited high sensitivity and specificity in predicting LUAD, with an area under the curve (AUC) value of 0.88 (P < 0.0001) (Fig. 5g). Additionally, Kaplan–Meier survival analysis showed that the median survival time in the high PLPP2-group was significantly lower than that of the low PLPP2-group (hazard response [HR] = 3.68, 95% confidence interval [CI] = 1.06–12.77, P = 0.04; Fig. 5h). Meanwhile, the multivariate Cox regression analysis showed that elevated PLPP2 was an independent prognostic risk factor for patients with LUAD (HR = 814, 95% CI = 1.51–438,194.06, P = 0.04; Fig. 5i). Furthermore, consistent outcomes were observed when conducting the prognostic analysis using the complete sample of 80 patients with follow-up data in Cohort 2 (Supplementary Fig. S5). Collectively, our findings unveiled that increased PLPP2 expression independently contributed to the prognostic risk assessment of early-stage LUAD patients.
Fig. 5
The elevated PLPP2 in LUAD served as an independent prognostic risk factor for LUAD. a PLPP2 mRNA levels in paired paracancerous and tumour tissues of patients in cohort 1 were tested by Q-PCR, n = 30. ****P < 0.0001; the student’s t test. b PLPP2 mRNA levels in BEAS-2B cells and LUAD cells were tested by Q-PCR, n = 3. Bars, SD; *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; the student’s t test. c The level of PLPP2 expressions in BEAS-2B cells and LUAD cells were tested by western blot, representative pictures were shown, n = 3. Bars, SD; ***P < 0.001, ****P < 0.0001; the student’s t test. d Representative images of IHC assay detecting levels of PLPP2 expression in paracancerous and tumour tissues of LUAD patients in cohort 2. e–f Statistic analysis of PLPP2 AOD values in paired paracancerous and tumour tissues of stage I LUAD patients in cohort 2, n = 42. Bars, SD; ****P < 0.0001; the student’s t test. g ROC analysis was used to evaluate the diagnostic accuracy of PLPP2 and calculate AUC value of stage I LUAD patients, n = 42. ****P < 0.0001. h 5-year survival probabilities of stage I LUAD patients in high and low PLPP2 groups were evaluated and showed by Kaplan–Meier curves, n = 42. *P < 0.05; Log-rank test. i Multivariate Cox hazard regression analysis of different clinical characters including age, gender and PLPP2 levels, n = 42
PLPP2-induced excessive proliferation of LUAD cells were dependent on lipid rafts formationTo investigate the role of PLPP2 in LUAD cells, we employed RNA-interference technology to downregulate PLPP2 expression in both A549 and NCI-H1299 cells. Immunofluorescence staining revealed a significant decrease in the localization of caveolin-1 and flotillin-1, which serve as lipid raft markers, on the cellular membrane (Fig. 6a-c). Lipid rafts played a crucial role in sensing extracellular signals that promote the malignant proliferation of tumor cells [31]. Therefore, CCK-8 assays were conducted to assess the malignant proliferation abilities of LUAD cells. Upon knockdown, both LUAD cell lines exhibited weakened proliferation capabilities (Fig. 6d-e). Similarly, the clonogenic growth assay demonstrated a significant reduction in the number of clonogenic colonies formed by LUAD cells following PLPP2 knockdown (Fig. 6f-g). Furthermore, depletion of PLPP2 also attenuated the resistance of LUAD cells to stress-induced apoptosis (Supplementary Fig. S6a-c). Additionally, transmission electron microscopy (TEM) images revealed that depletion of PLPP2 led to disruption within the inner membrane system, including compromised membrane integrity of endoplasmic reticulum and Golgi apparatus (Supplementary Fig. S6d). To further illustrate the potential of PLPP2 as a tumor marker, we employed a recombinant lentivirus to induce overexpression of PLPP2 in BEAS-2B and MLE-12 cells, which are normal lung epithelial cells. The findings revealed an elevation in caveolin-1 and flotillin-1 levels on the cell membrane (Fig. 6i-j) and enhanced cellular proliferation activity (Fig. 6k and l) subsequent to PLPP2 overexpression in normal lung cells. Furthermore, quantitative lipidomics analysis was conducted on PLPP2-overexpressed BEAS-2B cells utilizing the methodologies described in Fig. 4. Similarly, the differential metabolites were also enriched in signaling pathways associated with lipid rafts, such as cholesterol and glycerophospholipid metabolism. Furthermore, we observed a consistent pattern of alterations in the composition of lipid rafts, including SM (d18:0/22:0), SPH (d17:1), and SPH (d18:2) (Supplementary Fig. S7). Overall, our findings suggested that PLPP2 might enhance the survival of lung cancer cells by promoting proliferation and inhibiting cell death through enhancing lipid raft formation.
Fig. 6
Effects of PLPP2 on the content of lipid rafts and cell proliferation. a A549 and NCI-H1299 cells were fixed after transfected with target siRNAs and IF was performed with antibodies recognizing flotillin-1 (red) and caveolin-1 (green). b-c Statistic analyses of flotillin-1 and caveolin-1 AOD values in A549 and NCI-H1299 cells after transfected with target siRNAs, n = 3. Bars, SD; **P < 0.01, ***P < 0.001, ****P < 0.0001; one way ANOVA. d-e Effects of PLPP2 knockdown on cell proliferation by CCK-8 in A549 and NCI-H1299 cells, n = 3. Bars, SD; *P < 0.05, ***P < 0.001, ****P < 0.0001; one way ANOVA. f-g Effects of PLPP2 knockdown on cell proliferation by colony formation assays in A549 and NCI-H1299 cells, n = 3. Bars, SD; ****P < 0.0001; one way ANOVA. h BEAS-2B and MLE-12 cells were fixed after transfected with target lentivirus particles and IF was performed with antibodies recognizing flotillin-1 (red) and caveolin-1 (green). i-j Statistic analyses of flotillin-1 and caveolin-1 AOD values in BEAS-2B and MLE-12 cells after transfected with target lentivirus particles, n = 3. Bars, SD; ns: no significance, **P < 0.01, ****P < 0.0001; one way ANOVA. k-l Effects of PLPP2 OE on cell proliferation by CCK-8 in BEAS-2B and MLE-12 cells, n = 3. Bars, SD; *P < 0.05, **P < 0.01; one way ANOVA
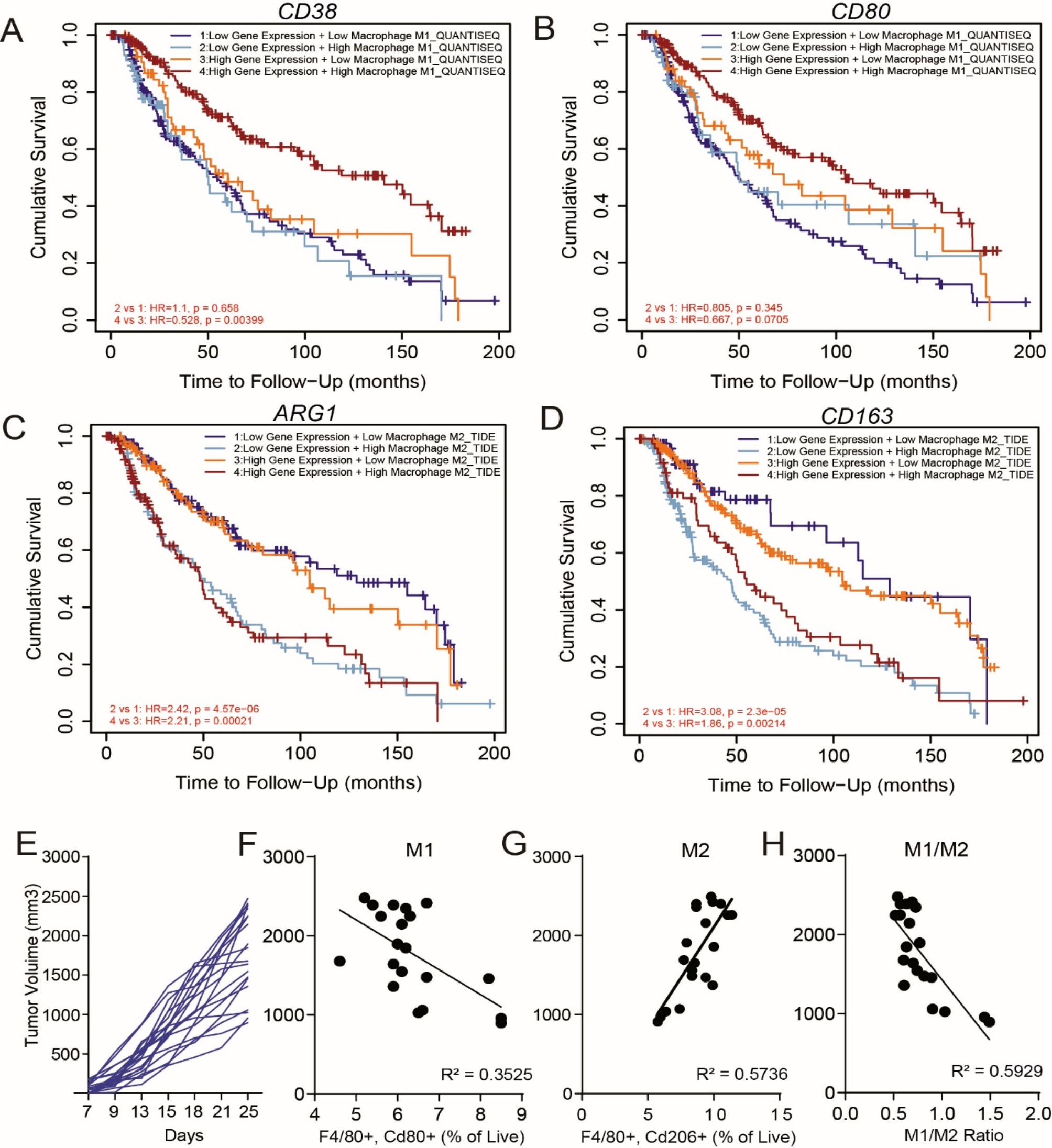
To further investigate the role of PLPP2 in promoting the malignant phenotype of LUAD cells through a lipid raft-dependent mechanism, we utilized a recombinant lentivirus to overexpress PLPP2 in human A549 and NCI-H1299 cells, as well as mouse LLC cells. Subsequently, we employed MβCD, a lipid raft-synthesis inhibitor, in subsequent experiments to reduce the abundance of lipid rafts on the membrane of LUAD cells [32]. We then assessed the impact of PLPP2 on the malignant phenotype of LUAD cells and observed that MβCD effectively inhibited excessive proliferation induced by PLPP2 overexpression (Fig. 7a and b). Similarly, application of a lipid raft inhibitor completely abrogated PLPP2-induced clonal overgrowth in LUAD cells (Fig. 7c and d). Immunofluorescence analysis revealed a significant increase in lipid raft contents in mouse LLC cells overexpressing PLPP2 (Fig. 7e and f). Next, the impact of PLPP2 on tumorigenesis was further examined in mice harboring subcutaneous tumors. As illustrated in Fig. 7g and h, overexpression of PLPP2 in LLC cells significantly enhanced tumor formation in mice. The group with PLPP2 overexpression exhibited notably larger tumor volume and largest tumor area compared to the vector-control group. However, treatment with MβCD effectively inhibited tumor growth in vivo, leading to significant reductions in both tumor volume and transverse largest area (Supplementary Fig. S8a and b). Immunohistochemistry (IHC) analysis was employed to evaluate lipid raft contents within the tumor tissue. Following PLPP2 overexpression, a marked increase was observed in caveolin-1 and flotillin-1 expression levels among mice bearing tumors; nevertheless, these effects were completely abrogated by the inhibitor of lipid raft (Supplementary Fig. S8c-e). Cell proliferation in tumor tissues was assessed by analyzing the percentage of Ki-67 and proliferating cell nuclear antigen (PCNA) positivity. MβCD significantly mitigated the overexpression of PLPP2-induced excessive proliferation, characterized by high-positive staining of Ki-67 and PCNA in tumor tissues (Supplementary Fig. S8f-h). Furthermore, there was a positive correlation between the levels of lipid rafts in tumor tissues and cell proliferation (Supplementary Figure S8i). These results from inhibition experiments further validated that PLPP2 might induce excessive proliferation of LUAD cells through its potential promotion of lipid raft formation.
Fig. 7
Inhibiting lipid raft synthesis in LUAD cells impeded the tumour-promoting effects of PLPP2. a-b Effects of PLPP2 OE on cell proliferation by CCK-8 in A549 and NCI-H1299 cells, n = 3. Bars, SD; *P < 0.05, **P < 0.01; two way ANOVA. c-d Effects of PLPP2 OE on cell proliferation by colony formation assays in A549 and NCI-H1299 cells, n = 3. Bars, SD; **P < 0.01, ***P < 0.001, ****P < 0.0001; two way ANOVA. e LLC cells were fixed after infected with target lentivirus particles and IF was performed with antibodies recognizing flotillin-1 (red) and caveolin-1 (green). f Statistic analyses of flotillin-1 and caveolin-1 AOD values in LLC cells after infected with target lentivirus particles, n = 3. Bars, SD; ***P < 0.001; the student’s t test. g Subcutaneous tumour images of LLC-vector group, LLC-PLPP2 OE group, LLC-vector plus MβCD treatment group and LLC-PLPP2 OE plus MβCD treatment group in C57BL/6 mice. h Tumour growth curves derived from target groups were shown, n = 6. Bars, SD; ***P < 0.001, ****P < 0.0001; two way ANOVA
Comments (0)