
Remember me


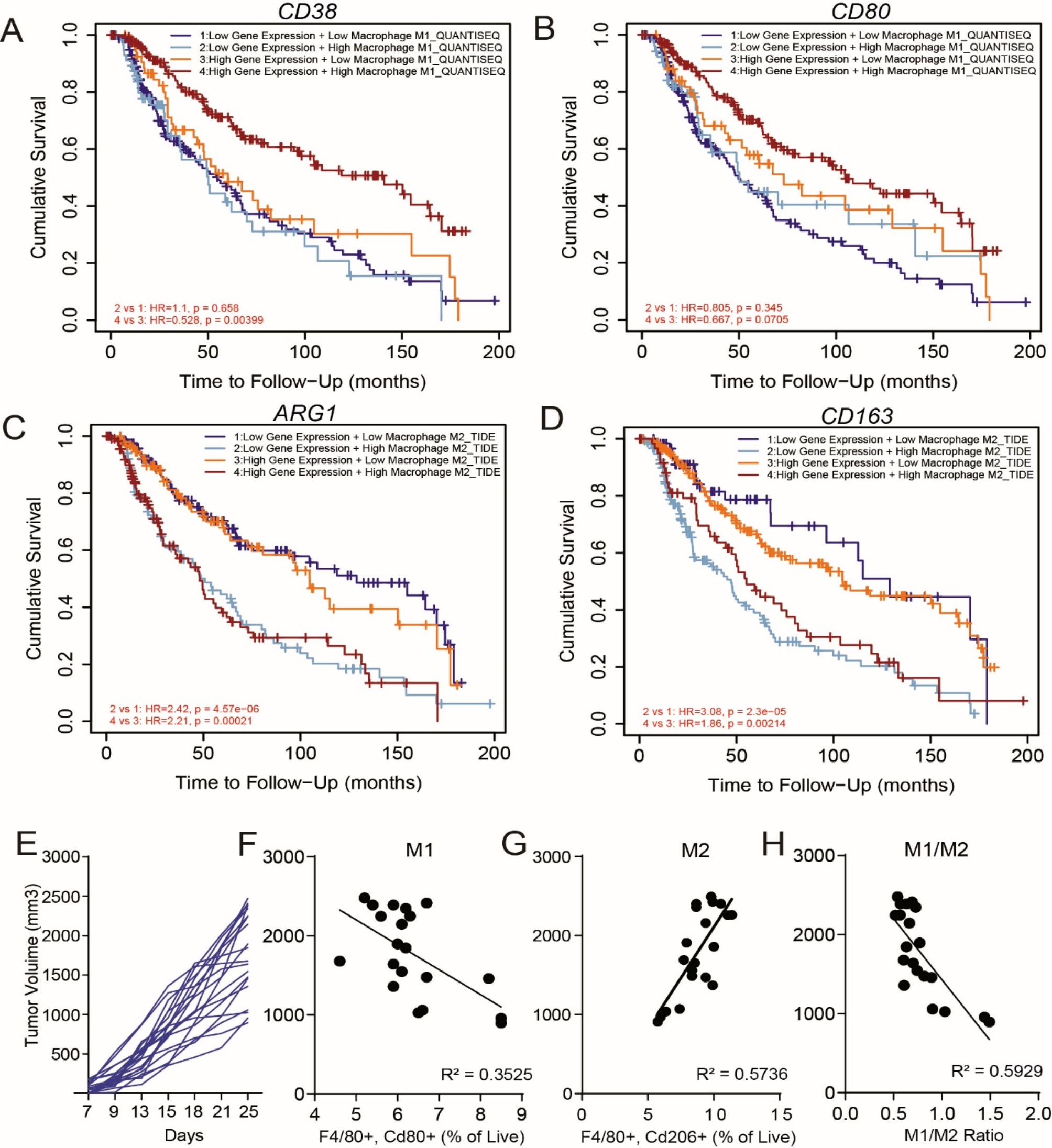


We established a chronic sleep deprivation model mouse to explore the effect of sleep disturbance on tumors. The results indicated that in the sleep-deprivated group, volume and weight of subcutaneous colon cancer tumors were increased (Fig. 1A-C). Additionally, more pulmonary nodules and increased infiltration were observed in the sleep-deprivated group compared to the regular sleep group in the metastasis mouse model (Fig. 1D, E).
Fig. 1
Sleep deprivation promotes occurrence and metastasis of CRC by GABA A, C Sleep deprivation increased the volume and weight of subcutaneous tumors. B The solid tumors were peeled off and weighted. C The volume of transplanted tumors was measured by the Vernier caliper every five days. D The number of tumors in the lung was counted in control group and SD group. E HE staining showed the tumors in the lung of mice. F ELISA assay showed the levels of GABA in the serum of SD group and control group mice. G The expression of GAD1, VGAT and GAT1 in cerebrum and cerebellum of mice was detected by western blot. H HE staining showed the primary tumors from AOM/DSS-induced colonic tumors in PBS group and GABA group. I The number of primary tumors in the colon was counted in PBS group and GABA group. J The expression level of E-cadherin, N-cadherin and Cyclin E was detected by IHC in primary tumors from respectively intraperitoneal injection PBS or GABA of AOM/DSS model mice. SD:sleep deprivation. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01.*P < 0.05
To further clarify the mechanisms linking sleep disorder and colon tumors, we assessed the levels of several sleep-related neurotransmitters in the serum of sleep deprivation mice, including GABA, NE, EPI, and 5-HT (Fig. 1F, S1A-C). Notably, the serum concentration of GABA significantly increased in the sleep-deprivated group. We examined brain proteins from the control and sleep-deprivated groups to determine the relationship between chronic sleep deprivation and GABA expression. The expressions of glutamate decarboxylase 1 (GAD1), GABA vesicular transporter (VGAT), and GABA transporter 1 (GAT1) in the cerebrum and cerebellum were examined by WB (Fig. 1G). The results showed that elevated expressions of GAD1 and VGAT in the sleep-deprivated group, indicating increased GABA synthesis and transport capacity in the brain tissue. Conversely, GAT1 expression was significantly decreased in the sleep-deprivated group. GAT1 can reuptake GABA in the synaptic space, which suggests that sleep deprivation maintains high GABA levels in the synaptic area, thus promoting GABA entry into the peripheral blood. We also observed the effect of GABA on the AOM-DSS mouse model (Fig. S1D). These results revealed that GABA increased the number of tumor lesions in the colon in mice (Fig. 1H-I, S1E). Furthermore, the immunohistochemical staining results suggested that GABA promoted proliferation and migration in primary colon tumors(Fig. 1J). In conclusion, our findings suggest that sleep deprivation promote colon cancer proliferation and migration by encouraging GABA synthesis and blood entry.
GABA inhibits ubiquitination of cMYC and promotes proliferation and migration of CRCPrevious studies have identified three kinds of GABA receptors: ionic GABAA, GABAC, and metabolic GABAB receptors. Type A and type B receptors have been widely studied and reported. We screened 30 pairs of colon cancer and paracancerous tissues through the GSE database. The heatmap showed it was the expression of the B receptor but not A receptor in colon cancer tissues that was significantly higher than in the paracancerous tissues (Fig. S2A). Then, we collected seven pairs of colon cancer tissues and detected the expression of GABARAP and GABBR2 in the tissues by qRT-PCR. The results showed that both GABARAP and GABBR2 were highly expressed in the tumor tissues, and the fold change of GABBR2 was more significant (Fig. S2B).
Therefore, we speculated that the B receptor were involved in regulating GABA in colon cancer. To further verify the mechanism of GABA-promoting colon cancer progression, we screened colon cancer cell lines with high expression of B receptors: SW480 and LoVo cells (Fig. S2C). We treated tumor cells with different concentrations of GABA in vitro and selected 100 μM for subsequent experiments (Fig. S2D). The results of CCK-8 assays (Fig. 2A), colony formation assay (Fig. 2B), wound-healing assays (Fig. 2C, D), and transwell assays (Fig. 2E) demonstrated that the proliferation ability and migration ability were enhanced when colon cancer cells were treated with GABA.
Fig. 2
GABA inhibits ubiquitination of cMYC and promotes proliferation and migration of CRC. A The proliferation of colon cancer cells was assessed via CCK-8 for 3 days. B The proliferation of colon cancer cells was assessed via colony formation assay for 10 days. C, D The wound healing assays showed that GABA significantly augmented the migrative ability of colon cancer cells. E The transwell assays indicated that GABA increased the migrative ability of colon cancer cells. F, H Treatment with GABA increased the volume and weight of subcutaneous tumors. G The solid tumors were peeled off and the weight was measured. H The volume of transplanted tumors was measured by the Vernier caliper per week. I, J The number of tumors in the lung was counted upon treated with GABA or PBS. K HE staining showed the tumors in the lung of mice. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01
The results of Western Blot analysis also showed that GABA treatment upregulated the expression of N-cadherin, vimentin, cyclin D1, and cyclin E while suppressing E-cadherin expression (Fig. S2E). In addition, we found increased volume and weight of subcutaneous xenografts in GABA-treated group compared with the PBS group (Fig. 2F-H). At the same time, the effect of GABA on tumor metastasis was verified by tail vein injection of colon tumor cells (Fig. 2I, H). HE staining indicated that the number and size of pulmonary metastatic nodules in the GABA-treated group were higher than the control group (Fig. 2K). These results confirmed that GABA is able to promote the proliferation and migration of colon tumors.
Studies have suggested that cMYC is abnormally expressed in about 70% of human tumors, regulating cell proliferation, differentiation, metabolism, and apoptosis [33]. We found that GABA treatment can upregulate the expression of cMYC in colon cancer cells. Ubiquitination of cMYC protein is considered as an essential means of targeting cMYC to inhibit tumors and has been widely studied [34]. Therefore, we examined whether GABA inhibited the ubiquitination of cMYC protein. Cycloheximide (CHX), a protein synthesis inhibitor, was used to explore the degradation of cMYC protein. After treatment with CHX, the degradation rate of cMYC protein in GABA group was significantly lower than that in PBS group, indicating that GABA may increase the stability of cMYC protein (Fig. 3A). In addition, MG132, a proteasome inhibitor, take an important role influencing the effect of GABA on the deubiquitination of cMYC protein (Fig. 3B), which affirmed that the inhibition of cMYC degradation by GABA is dependent on protease. We further examined the ubiquitinated form of endogenous cMYC. The cMYC protein was immunoprecipitated with an anti-cMYC antibody, and the ubiquitinated condition of cMYC was detected with an anti-ubiquitinated antibody. In the presence of MG132, the ubiquitinated form of endogenous cMYC in the GABA treatment group was significantly lower than in the PBS group (Fig. 3C). It suggests that GABA treatment reduced the ubiquitination form of cMYC in colon cancer cells and enhanced the stability of cmyc protein. To further explore the effect of GABA treatment on promoting the proliferation and migration of colon cancer cells through regulating cMYC, we knocked down cMYC in GABA-stimulated colon cancer cells. The results revealed that cMYC knockdown weakened the regulation of GABA on the proliferation and migration of colon cancer cells (Fig. 3D-G, S2F).
Fig. 3
GABA inhibits ubiquitination of c-MYC and promotes proliferation and migration of CRC. A Colon cancer cells were treated with GABA followed by treatment with cycloheximide (CHX) for the indicated times. The intensity of cMYC expression at each time point was quantified by densitometry and plotted against time. B The expression of cMYC in colon cancer cells treated with GABA for 3 days and then incubated with or without MG132 for 6 h via western blot. C Ubiqutin assays of colon cancer cells treated with GABA and MG132. D The western blot analysis revealed that cMYC knockdown reversed the effects of GABA-induced promotion of proliferation and migration in colon cancer cells. E CCK-8 assays showed that cMYC knockdown reverses the effects of GABA-induced promotion of proliferation in colon cancer cells. F Colony formation assays revealed that cMYC knockdown reverses the effects of GABA-induced promotion of proliferation in colon cancer cells. G Transwell assays revealed that cMYC knockdown reverses the effects of GABA-induced promotion of migration in colon cancer cells. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01.*P < 0.05
In summary, our data suggest that GABA enhances the stability of cMYC protein, inhibits its ubiquitination degradation, and promotes colon cancer cell proliferation and migration.
GABA promotes the recruitment of macrophages by CRC and induces M2 polarization through exosomesIt has been previously reported that B-cell-derived GABA could affect the classification of macrophages and inhibit anti-tumor immunity, and another study found that GABA secreted by tumor cells were to activate GSK3 β-Pathway to inhibit CD8+T cell-mediated tumor immunity [11, 12]. What is mentioned above suggests that GABA is probably involved in the mutual regulation of tumor immunity. We performed immunofluorescence to detect macrophage markers in mouse lung metastases. Interestingly, macrophage infiltration around lung metastases was increased in GABA-treated group mice. (Fig. 4A). Additionally, co-culturing with GABA-induced colon cancer cells increased the migration ability of macrophages (Fig. S4A). It suggests that GABA promotes the recruitment of colon cancer cells to peripheral macrophages, which is related to tumor progression.
Fig. 4
GABA promotes the recruitment of macrophages by CRC and induces M2 polarization through exosomes. A Immunofluorescence images showed that macrophages were relatively scarce in lung metastases of PBS group, while were abundant in GABA group. B The expression of CD206, CD163, Arginase1 and CD86 of THP1 cells upon administration of GABA were detected by Western blot and qRT-PCR. C. The expression of CD206, CD163, Arginase1 and CD86 of THP1 cells co-cultured with GABA-induced colon cancer cells or not by Western blot. D The expression of CD206, CD163, Arginase1 and CD86 of THP1 cells co-cultured with GABA-induced SW480 or not was detected by qRT-PCR. E, F Western blot and qRT-PCR showed that the expression of M2 makers (CD206, CD163, Arginase1) in THP1 cells was increased more significantly when added GABA-induced SW480-CM by qRT-PCR, while M1 maker was decreased. G Representative immunofluorescence image showed the internalization of PKH26-labeled LoVo-derived exosomes by THP-1 cells. H Western blot showed that the expression of M2 makers in THP1 cells was increased more significantly when added GABA-induced-SW480 derived exosomes, while M1 maker was decreased. I Immunofluorescence image showed that THP1 cells exhibited more pronounced M2 polarization when incubated GABA-induced SW480-derived exosomes for 3 days. J Western blot showed that GW4869(an inhibitor of exosome secretion) reversed the strengthened M2 polarization of THP1 cells upon incubated GABA-induced SW480-derived exosomes. SW480P: PBS-induced SW480 cells. SW480G:GABA-induced SW480 cells. LoVoP: PBS-induced LoVo. LoVoG:GABA-induced LoVo cells. Exo: Exosomes.CM: conditioned medium. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01.*P < 0.05. ns means the difference was not significant
TAMs are the primary members of immune cells infiltrating into tumors. M2 macrophages play an irreplaceable role in regulating tumor growth and metastasis. Here, we investigated the polarization of macrophages recruited in the tumor microenvironment. We induced THP1 cells with PMA to form M0 macrophages, and confirmed its morphological changes (Fig. S3B). The augment of M0 marker CD68 was verified by qRT-PCR (Fig. S3C). First, we found that the polarization of THP1 cells treated with GABA alone did not change significantly (Fig. 4B). However, M2 polarization was ensured after co-culturing with normal colon cancer cells through WB and qRT-PCR. Intriguingly, when co-cultured with GABA-induced colon cancer cells, it exhibited more pronounced M2 polarization (Fig. 4C, D, S3D). Combined with previous experimental results, we speculated that GABA could promote the interaction between colon cancer cells and macrophages. We further detected the effect of the conditioned medium of colon cancer cells on macrophage polarization, and the results showed that GABA-treated colon cancer cell medium induced M2 polarization of macrophages more significantly (Fig. 4E, F, S3E, F).
It has been reported that tumor cells can secrete numerous exosomes, which take a great part in cell interaction. We speculated that GABA may promote the secretion of exosomes by colon cancer cells, thereby causing M2 polarization of macrophages. We collected the culture medium of colon cancer cells in the control and GABA treatment groups respectively to extract exosomes, further identified the extracted exosomes (Fig. S 3G- I). Exosomes were labeled with the fluorescent dye PKH26, and our data showed that exosomes derived from colon cancer cells were fused to macrophages (Fig. 4G). The extracted exosomes of colon cancer cells were co-cultured with THP1 cells for 48 h and detected the polarization of macrophages via WB. These results revealed that the M2 polarization of macrophages induced by exosomes secreted by GABA-induced colon cancer cells was more prominent (Fig. 4H). Immunofluorescence showed the same results (Fig. 4I, S3I). In addition, GW4869, an exosome inhibitor, can reverse this M2 polarization effect of tumor cells on macrophages (Fig. 4J, S3J, K).
In conclusion, our data demonstrates that GABA-induced colon cancer cells recruit macrophages in the microenvironment and aggravate the M2 polarization-promoting effect of tumor cells on macrophages through the exosome pathway rather than direct impact of GABA on macrophages.
Exosome miR-223-3p promotes M2 polarization of macrophages, which aggravates the proliferation and migration of CRCMiRNAs are the most abundant substances in exosomes and participates in regulating various cells. To explore the mechanism of GABA-induced colon cancer cell-derived exosome promotion of M2 polarization in macrophages, miRNA sequencing in exosomes was used to perform large-scale expression profiling(Supplementary Table 1). We screened nine up-regulated miRNAs and five down-regulated miRNAs (Fig. 5A) and verified the sequencing results via qRT-PCR (Fig. S4A). Reports have indicated that mir-150-5p and miR-223-3p are involved in the regulation of macrophage M2 polarization. MiR-150 is an immune-related miRNA, which is upregulated in various cancers [35]. MiR-223 is involved in the regulation of inflammation, tumor proliferation, and invasion after transcription. Zhuang et al. reported that miR-223-3p, a new regulator of macrophages, could inhibit standard M1 polarization and promote M2 polarization [36]. More importantly, miR-223-3p acts as an intermediate communication signal between tumor and immune cells in TME [18]. Therefore, we further detected the content of miR-223-3p and miR-150-5p in exosomes. QRT-PCR results demonstrated that GABA promoted the inclusion of miR-223-3p in exosomes, and the content of miR-223-3p in the medium decreased significantly, while the range of miR-150-5p did not change significantly (Fig. S4B). Additionally, treatment with RNAase A and Triton X-100 significantly reversed the increase of miR-223-3p expression in GABA-induced exosomes, however, RNAase alone had no significant effect (Fig. 5B).
Fig. 5
Exosome miR-223-3p promotes M2 polarization of macrophages, which aggravates the proliferation and migration of CRC. A A cluster heatmap of the expression profiles of miRNAs in the exosomes derived from PBS-induced LoVo cells and GABA-induced LoVo cells. B The qRT-PCR analysis of miR-223-3p in the CM of LoVo cells was treated with RNase A (3U/μg) alone or combined with Triton X-100 (0.1%) for 20 min. C-D The expression of CD206, CD163 and CD86 in THP1 cells upon miR-223-3p overexpression or not by qRT-PCR and Western blot. E Immunofluorescence assay indicated that miR-223-3p overexpression increased the expression of M2 makers in THP1 cells. F The proliferation of SW480 cells co-cultured with THP1 overexpressed miR-223-3p or not was assessed via CCK-8 for 3 days. G Proliferation of SW480 cells co-cultured with THP1 cells overexpressed miR-223-3p or not was assessed via colony formation assay for 10 days. H The transwell assays indicated that co-cultured with THP1 cells overexpressed miR-223-3p increased the migrative ability of SW480 cells. I The wound healing assays revealed that co-cultured with THP1 cells overexpressed miR-223-3p increased the migrative ability of SW480 cells. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01.*P < 0.05. ns means the difference was not significant
After overexpression of miR-223-3p in macrophages, qRT-PCR, WB, and immunofluorescence results showed that the expression of the M2 polarization marker significantly increased (Fig. 5C-E). We found that miRNAs were correlated with the MAPK pathway via KEGG analysis (Fig. S4C). The activation of the MAPK pathway has been proven to be linkedwith the M2 polarization of macrophages. P38 inhibitor and ERK inhibitor were used in THP1 cells overexpressing miR-223-3p, with the results showing that inhibition of the ERK pathway could effectively reverse the M2 polarization of macrophages induced by miR-223-3p. In contrast, inhibition of the p38 pathway had no significant effect (Fig. S4D). We further co-cultured macrophages overexpressing miR-223-3p with colon cancer cells to observe the feedback effect of macrophages on colon cancer. The results proved that co-cultured with mimic transfected macrophages could significantly improve the proliferation and migration of tumor cells(Fig. 5F-I, S4E-H). Furthermore, the results of subcutaneous tumor implantation in mice indicated that co-culturing with macrophages transfected with mimic could accelerate the growth of colon cancer cells (Fig. S5A-C). The results of lung metastasis of tail vein tumor also showed that co-culturing with mimics transfected macrophages enhanced the migration ability of colon cancer cells (Fig. S5D-F).WB revealed that the expressions of increased N-cadherin, vimentin, cyclin E and cyclin D, decreased E-cadherin, in colon cancer cells co-cultured with THP1 cells transfected with mimic (Fig. S5G).
We also collected mimic transfected macrophage-conditioned medium for culturing colon cancer cells. Results showed that the medium of macrophages transfected with mimics significantly enhanced the proliferation and migration of colon cancer cells (Fig. 6A-C, S5H). WB also revealed the same results (Fig. S 5I). We further measured the level of cytokines in a macrophage culture medium with a cytokine Chip (Fig. 6D). At the same time, qRT-PCR was performed to verify the mRNA level of cytokines, and we found that IL-17 increased most significantly (Fig. 6E). The expression of IL-17 increased or decreased upon miR-223-3p overexpression or inhibition in macrophages (Fig. 6G). IL-17 is a typical pro-inflammatory cytokine, which plays a regulatory role in host defense, tissue repair, inflammatory immunity and tumor progression [37]. Moreover, IL-17 augmented the expression of vimentin, CyclinD1, and cyclinE, while attenuating the expression of E-cadherin in colon cancer cells (Fig. 6H). IL-17 was reported to activate JAK-STAT3 and NF- κ B, and other signaling pathways [38]. It can also promote the expression of PD-L1 and cause tumor immune escape [39]. Isibizumab was used to block the IL-17 receptor in colon cancer cells. The results of WB analysis suggested that the JAK-STAT3 pathway were inhibited, and the expression of the marker proteins of proliferation and migration were also reversed (Fig. 6F).
Fig. 6
Exosome miR-223-3p promotes M2 polarization of macrophages, which aggravates the proliferation and migration of CRC. A CCK-8 assays showed that proliferation of colon cancer cells was significantly augmented when added the CM of THP1 cells overexpressed miR-223-3p. B The transwell assays indicated that migrative ability was significantly augmented when added the CM of THP1 cells overexpressed miR-223-3p. C The colony formation assay indicated that migrative ability was significantly augmented when added the CM of THP1 cells overexpressed miR-223-3p. D A cluster heatmap of the expression profiles of cytokines in the CM of THP1 cells upon overexpression miR-223-3p or not via ELISA assays. E The expression of cytokines in THP1 cells upon overexpression miR-223-3p or not was detected by qRT-PCR. F Western blot indicated that inhibition of IL17 blocked the altered expression of E-cadherin, N-cadherin, Vimentin, Cyclin E and Cyclin D1 in colon cancer cells co-cultured with THP1 cells overexpressed miR-223-3p. G Western blot showed that the expression of IL17 was conspicuously increased or decreased in THP1 cells upon miR-223-3p overexpression or knockdown in THP1 cells, respectively. H The expression of E-cadherin, Vimentin, Cyclin E and Cyclin D1 in colon cancer cells upon administration of IL17 were detected by Western blot. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01.*P < 0.05. ns means the difference was not significant
In conclusion, our data suggested that tumor-derived exosomes miR-223-3p activates the MAPK pathway of macrophages to cause M2 polarization and secrete IL-17 to aggravate the proliferation and migration of colon cancer.
GABA promotes the proliferation and migration of CRC by inhibiting ubiquitination of cMYC through miR-223-3pAlthough miR-223-3p is a conservative anti-inflammatory miRNA, studies have indicated its function in tumor proliferation and invasion in prostate cancer, breast cancer, and other tumors [22, 40]. Therefore, we further explored whether the endogenous expression of miR-223-3p in colon cancer cells impacted on tumor proliferation and migration. Results showed that overexpression of miR-223-3p can promote the proliferation of colon cancer cells (Fig. S6A, B), while increasing the migration ability of colon cancer cells (Fig. S6C, D). Our previous data proved that GABA could increase the stability of cMYC protein in tumor cells and reduce ubiquitination degradation. Notably, after treating with CHX in miR-mimics transfected colon cancer cells, the degradation rate of cMYC was also significantly slower than that of the control group (Fig. 7A). Immunoprecipitation also showed that the overexpression of miR-223-3p could reduce the ubiquitinated form of cMYC in tumor cells (Fig. 7B). This suggests that overexpression of miR-223-3p inhibits the normal ubiquitination and degradation process of cMYC protein, resulting in the relative increase of cMYC protein expression in tumor cells.
Fig. 7
GABA promotes the proliferation and migration of CRC by inhibiting ubiquitination of cMYC through miR-223-3p. A Colon cancer cells were overexpressed miR-223-3p followed by treatment with cycloheximide (CHX) for the indicated times. The intensity of cMYC expression at each time point was quantified by densitometry and plotted against time. B Ubiqutin assays of colon cancer cells transfected with miR-mimic followed by treatment with MG132. C. CCK-8 assays revealed that knockdown miR-223-3p reversed the GABA-induced promotion of proliferation in colon cancer cells. D The colony formation assay indicated that knockdown miR-223-3p reversed the GABA-induced promotion of proliferation in colon cancer cells. E The transwell assays showed that knockdown miR-223-3p reversed the GABA-induced strengthened migrative ability of colon cancer cells. F Western blot showed that inhibition of miR-223-3p reversed the GABA-induced promotion of proliferation and migration in colon cancer cells. G Ubiqutin assays indicated that inhibition of miR-223-3p blocked the altered ubiquitination of cMYC in colon cancer induced by GABA. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01.*P < 0.05
In this study, we intended to investigate whether GABA promoted the proliferation and migration of colon cancer by regulating the ubiquitination of cMYC protein through miR-223-3p. Inhibition of miR-223-3p can significantly reverse the promotion of GABA on the increase and migration of SW480 and LoVo cells (Fig. 7C-F). GABA inhibited the ubiquitination and degradation of cMYC protein while inhibition of miR-223-3p reversed the inhibitory effect of GABA on cMYC ubiquitination (Fig. 7G).
In conclusion, our data suggest that GABA promotes the proliferation and migration of colon cancer cells by stimulating the endogenous expression of miR-223-3p and the stability of cMYC proteins.
CMYC can reverse regulate the expression of miR-223-3pCoincidently, we found that the expression of miR-223-3p increased or decreased upon cMYC overexpression or inhibition in colon cancer cells (Fig. S6E, F). As a transcription factor, cMYC is involved in the regulation of a variety of tumors. Therefore, cMYC may acts as a transcription factor to regulate the transcription of miR-223-3p.
Data from the Jaspar database indicated a potential binding site for cMYC in the miR-223-3p promoter (Fig. S6G). Chip assays showed that compared with IgG-bound samples, cMYC-bound complexes were significantly enriched in the promoter region of miR-223-3p (Fig. S6H).
In conclusion, these data suggest that cMYC regulates miR-223-3p reversely.
Mir-223-3p targets E3 ligase CBLB to regulate ubiquitination of cMYCTo further clarify the specific mechanism of miR-223-3p regulating cMYC ubiquitination, we screened the target genes of miRNA from miRDB, miRWalk and TargetScan databases respectively (Fig. 8A). Notably, there are 56 downstream target genes in the intersection, three of which related to ubiquitination regulation, namely FBXW7, CBLB and SIAH1. QRT-PCR and WB results demonstrated that CBLB differentiates under the intervention of mimic and inhibitor, while both FBXW7 and SIAH1 performed poorly (Fig. 8B, C). Together, we also found that miR-223-3p may bind onto the 3'UTR region of the mRNA of CBLB (Fig. 8D). To confirm that CBLB was a target of miR-223-3p, 293 T cells were transfected with dual luciferase reporter plasmid containing wild-type CBLB 3'UTR (WT) or mutated type (MUT), followed by transfection with miR-223-3p mimic (miR-mimic) or negative control (mimic-NC). The results showed that the luciferase activity in 293 T cells was reduced upon the overexpression of miR-223-3p in the WT group but not in the MUT group (Fig. 8E).
Fig. 8
Mir-223-3p targets E3 ligase CBLB to regulate ubiquitination of cMYC. A Venn diagram indicated that the overlap of downstream genes of miR-223-3p predicted by miRDB, miRWalk and TargetScan database. B-C qRT-PCR and western blot showed that the expression of CBLB was conspicuously decreased or increased upon miR-223-3p overexpression or knockdown respectively in LoVo cells, while the difference of SIAH1 and FBXW7 was not significant. D A miR-223-3p binding site on the 3'UTR of the CBLB mRNA was predicted by TargetScan database. E The binding of miR-223-3p on CBLB 3’UTR was evaluated by dual luciferase reporter assay.293 T cells were transfected with reporter plasmid containing wild type CBLB 3'UTR(WT) or mutant type (MUT) respectively, followed by transfection with mir-223-3p mimic (miR-mimic) or negative control (miR-NC). F Western blot showed that the expression of proliferation and migration markers and cMYC was significantly decreased or increased in colon cancer cells upon CBLB overexpression or knockdown in colon cancer cells. G Colon cancer cells were overexpressed CBLB followed by treatment with cycloheximide (CHX) for the indicated times. The intensity of cMYC expression at each time point was quantified by densitometry and plotted against time. H Ubiqutin assays of colon cancer cells transfected Sh-CBLB followed by treatment with MG132. I CO-IP and Western blot showed that endogenous cMYC and CBLB bind to each other. J Immunofluorescence images showed colocalization of CBLB and cMYC in colon cancer cells. All data were revealed as mean ± standard deviation (SD) for no less than three independent experiments. Significant P values showed as ***P < 0.001.**P < 0.01. ns means the difference was not significant
Following, we verified the function of CBLB in colon cancer. CBLB is a proto-oncogene considered as a new target for tumor immunotherapy [41]. However, its expression in colon cancer has not yet been reported. We collected colon tissue samples from 10 patients with colon cancer. WB analysis results showed CBLB was highly expressed in the non-cancerous tissue of colon but lower in the tumor tissues (Fig. S7A). Immunohistochemistry also showed the same results (Fig. S 7B). We also detected the expression of CBLB in subcutaneous colon cancer tumors of control group and SD group. WB analysis results showed that sleep-deprivation decreased the expression of CBLB in subcutaneous colon cancer tumors, while cMYC was highly expressed in subcutaneous colon cancer tumors of SD group (Fig. S 7C). We further transfected SW480 and LoVo cells with CBLB overexpression plasmid and knockdown lentivirus. We found that knockdown of CBLB enhanced the proliferation and migration of tumor cells, while overexpression had the opposite effect (Fig. S7D-F, S8A-C). WB analysis results revealed that knockdown of CBLB increased the expression of cMYC and promoted the expression of proliferation and migration marker proteins, while overexpression of CBLB led to the opposite effect (Fig. 8F). Immunofluorescence results also indicated that downregulating the expression of CBLB promoted the proliferation and migration of colon cancer cells (Fig. S9A). Whereas, the results of subcutaneous tumor formation in mice also revealed that the expression of CBLB was negatively correlated with tumor proliferation (Fig. S9B-D).
We further explored the specific mechanism of cMYC down-regulation induced by CBLB with CHX to detect the degradation of cMYC protein. It was found that the degradation rate of the cMYC protein was significantly faster when CBLB was overexpressed in colon cancer cells (Fig. 8G), compared to the cMYC protein, which was considerably slower when CBLB was downregulated (Fig. S9E). This indicated that CBLB reduced the stability of the cMYC protein. In addition, GABA-induced colon cancer cells were treated with MG132, resulting in increased cMYC expression. However, MG132 did not significantly increase the expression of cMYC in CBLB knockdown colon cancer cells. While MG132 treating CBLB overexpressed colon cancer cells, it showed consistent results (Fig. S 9F, G). It indicates that CBLB regulates the degradation of cMYC through a proteasome-dependent pathway. We further explored the polyubiquitinated form of endogenous cMYC in CBLB knockdown colon cancer cells. In the presence of MG132, the ubiquitination form of endogenous cMYC protein in CBLB knockdown group was significantly reduced (Fig. 8H). To investigate further mechanism CBLB regulates cMYC protein levels, we explored whether CBLB directly binds to cMYC. Endogenous immunoprecipitation demonstrated that CBLB protein binds to cMYC (Fig. 8I, S9H). Immunofluorescence analysis of SW480 and LoVo cells revealed colocalization of CBLB and cMYC in cells (Fig. 8J). These results revealed that CBLB, as an E3 ligase, can recognize and bind the substrate cMYC.
Comments (0)