
Remember me



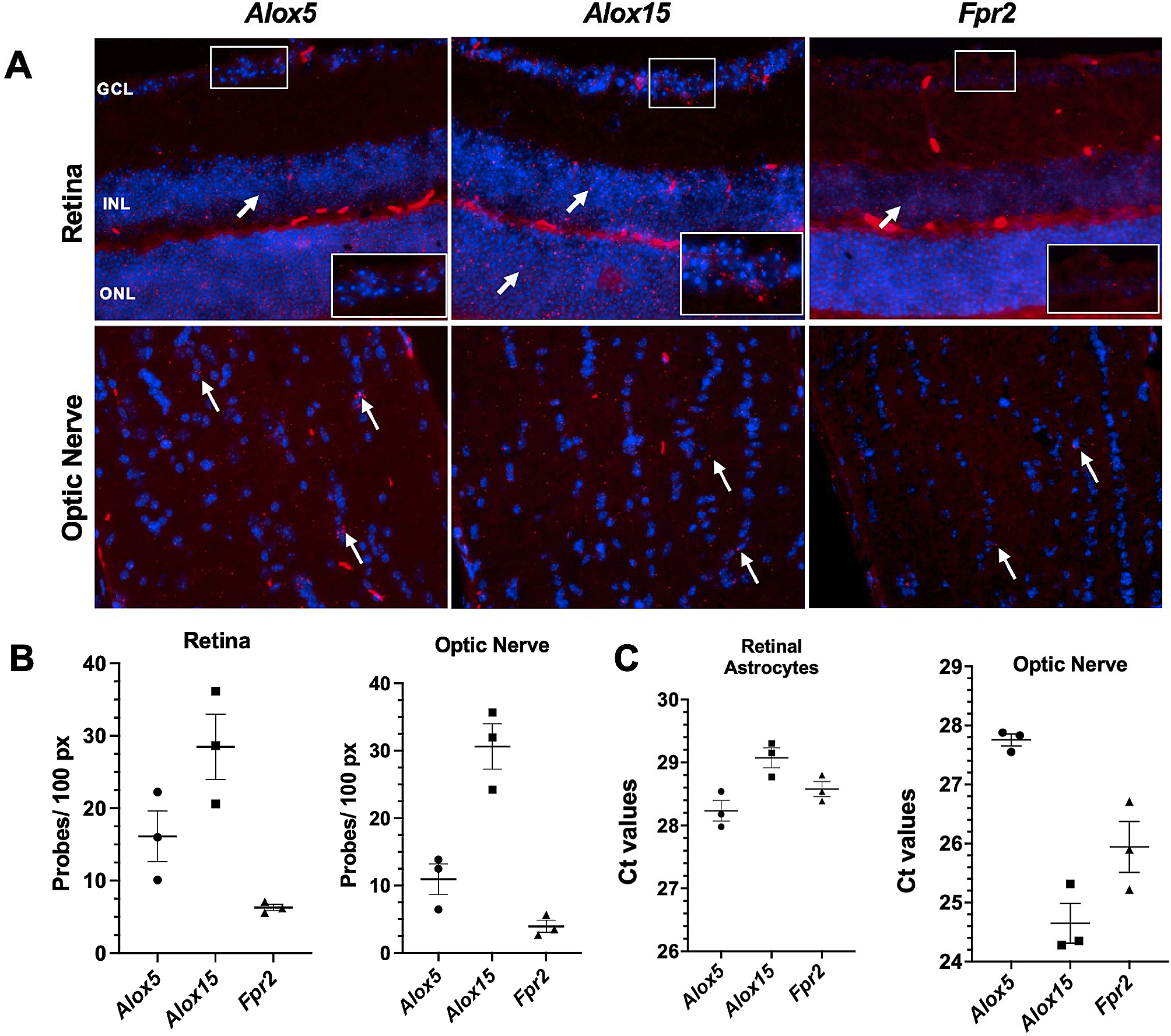
To better understand the progression of IDHmut astrocytomas to grade 4, we generated and integrated matched WGS and strand-specific RNA-seq data from five Tampere Glioma cohort (TG) patients (TG01–TG05) as well as RNA-seq data from TG06 (Fig. 1a). The key findings were validated using a larger in-house targeted sequencing cohort of 25 patients and published data from TCGA, the ICGC, and the GLASS consortia.
Fig. 1
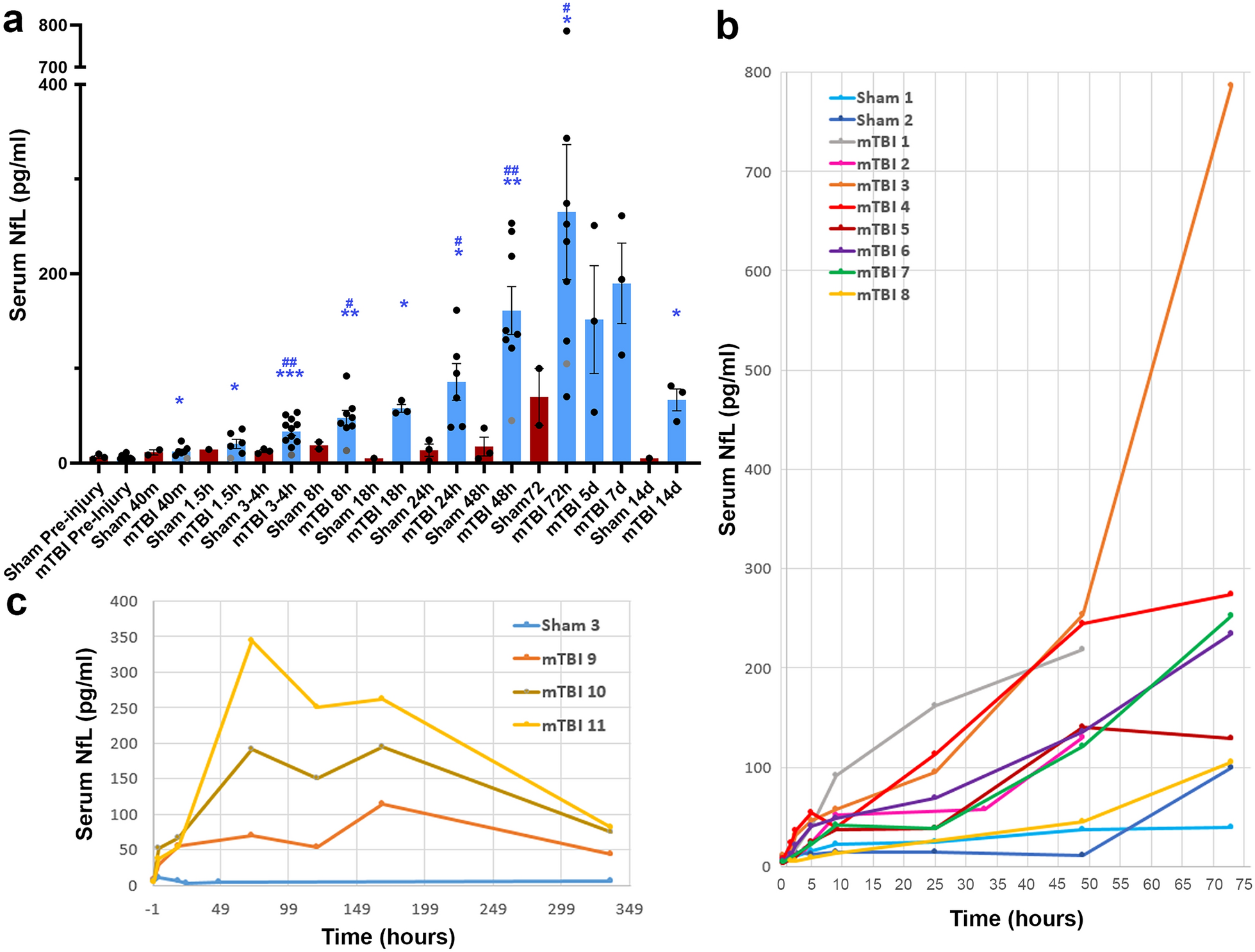
Progression to grade 4 is associated with increased hypoxia response, proliferation, and DNA repair. a The discovery cohort comprises six patients with IDHmut low-grade gliomas and relapsed grade 4 tumors. Cases TG01–TG05 were analyzed with WGS and TG01–TG06 with RNA-seq. A larger in-house IDHmut astrocytoma cohort was subjected to targeted DNA-sequencing for validation. In addition, 59 TCGA-ICGC WGS samples, 595 TCGA diffuse glioma RNA-seq samples, and 79 matched GLASS IDHmut astrocytoma samples were used as public validation data cohorts. SNV: single nucleotide variant. b Clinical courses of the patients in the discovery cohort. All six patients received radiation treatment and most patients received chemotherapy, but not TMZ, before progressing to grade 4. Vertical black lines represent surgeries and resected samples shown in gray were not measured as part of the discovery cohort. A: astrocytoma, OA: oligoastrocytoma. Tumor type is followed by the tumor grade. c Representative H&E images of the tumors in the discovery cohort with scale bars corresponding to 250 µm (low magnification) and 50 µm (high magnification). d More protein-altering mutations were detected in TG01 than in other cases (TG02–TG05) in the WGS data. e TG01 samples with full MSH2 deletion and DNMT3A inactivation showed a clear CG > TG substitution signature. f–g GO enrichment analysis for upregulated (f) and downregulated (g) genes. Upregulated genes were related to DNA repair, cell proliferation, and angiogenesis, whereas downregulated genes were associated with GPCR signaling pathways and the regulation of development. h Hypoxia response, cell cycle G2/M-phase, and S-phase gene set Z-score activities were upregulated in grade 4 tumors, except for TG01b, where proliferation activities slightly decreased. Gene set activities were scaled between 0 and 1 for visualization
Clinical course and pathological evaluation of the cases in the discovery cohortAll discovery cohort patients received a primary tumor diagnosis at the age of 25–35 years, except TG01 (age group 18–20) (Fig. 1b, Additional file 2: Fig. S1a and Supplementary Text, Additional file 1: Table S1). Most primary tumors were grade 2 IDHmut astrocytomas; TG02 was initially diagnosed as a grade 3 oligoastrocytoma and TG04 as a grade 3 astrocytoma (Fig. 1b–c, Additional file 2: Fig. S1a–b, Additional file 1: Table S1). All tumors eventually progressed to grade 4 and were at the time diagnosed as secondary glioblastomas. Their diagnosis was later updated to grade 4 astrocytoma based on the WHO 2021 guidelines [3]. Importantly, all patients received radiation during the course of the disease. TG05 was irradiated seven years before the first relapse (due to tumor regrowth based on magnetic resonance imaging (MRI) of the brain), after which chemotherapy alone was administered (Fig. 1b, Additional file 2: Fig. S1a). TG04 was the only patient who received radiation alone before disease progression. For TG01 and TG03, radiation was an initial postoperative treatment (therapy administered between surgeries), followed by chemotherapy closer to the progression than previous surgery. After the initial diagnosis, consecutive cycles of radiation and chemotherapy were administered to patients TG02 and TG06. Before progression to grade 4, patients TG01, TG03, and TG06 received PCV (procarbazine, lomustine, and vincristine) chemotherapy, whereas patients TG02 and TG05 received CCNU (lomustine) chemotherapy. Notably, none of the patients received TMZ before disease progression as it was not part of the standard of care at the time of operation. In surgeries for sequenced grade 2–3 tumors, a residual tumor mass was detected in TG04a and TG05a (Additional file 1: Table S1, Additional file 2: Supplementary Text).
For each of these cases, we analyzed matched tumors before and after progression to grade 4 (Fig. 1b and Additional file 2: Fig. S1a). All tumors harbored a missense mutation in IDH1 at p.R132 (p.R132G in TG01 and p.R132H in TG02–TG06) (Additional file 1: Table S1), and sample TG05b additionally had a copy-neutral loss-of-heterozygosity in IDH1 p.R132H. Inactivating TP53 and ATRX alterations were detected in most cases. Progressed grade 4 tumors inherited these driver mutations, consistent with previous reports [24]. No 1p19q co-deletions or TERT promoter, CIC gene, or FUBP1 mutations were detected.
Based on DNA methylation analysis, the grade 4 samples belonged to the ‘high-grade astrocytoma’ (A IDH, HG) methylation class (Additional file 2: Fig. S1c and Additional file 1: Table S1) [12]. Many tumors in this class progressed from low-grade astrocytomas. TG01b, TG02b, TG03b, and TG04b had high-confidence classification scores (> 0.84) [25]. TG05b had a score below 0.5 and should generally be discarded if high tumor cell content is assumed [25]; however, it clustered nicely with other A IDH, HG samples in the tSNE plot. To conclude, the samples in our cohort clearly represent IDHmut diffuse astrocytomas.
MSH2 and DNMT3A inactivation in hypermutator case TG01To estimate the mutation load in the samples, we counted the protein-altering mutations (excluding known germline variants) in the WGS data. Mutation counts increased, but only moderately, in grade 4 tumors. The total number of coding mutations ranged between 22 and 85 in all tumors except the hypermutator TG01 tumor, which harbored 134 nonsynonymous mutations before progression and 189 mutations in the grade 4 tumor (Fig. 1d and Additional file 1: Table S3). Interestingly, the TG01 sample exhibited a distinct mutational pattern characterized by C > T substitutions (Fig. 1e and Additional file 2: Fig. S1d–e), representing a mixture of SBS1 and SBS15 mutational signatures. SBS1 is observed in many human cancers and is positively correlated with patient age, whereas SBS15 is associated with defective DNA mismatch repair (MMR) and microsatellite instability [20, 26, 27]. The tumors of TG01 harbored inactivating alterations in MSH2 and DNMT3A. MSH2, which encodes a member of the DNA MMR complex, was homozygously deleted and markedly downregulated in the TG01 tumor (Additional file 2: Fig. S1f). Furthermore, de novo DNA methyltransferase DNMT3A was affected by a p.P904L (chr2:25,234,307:G > A) mutation, disrupting its methyltransferase domain, and loss-of-heterozygosity caused by a large 40 Mb deletion in chromosome 2 (Additional file 1: Table S3). The expression of DNMT3A was lower in TG01 than in other cases (Additional file 2: Fig. S1f). DNMT3A p.P904 mutations are rather common and located in the catalytic domain, leading to decreased protein stability [28,29,30], suggesting defective DNMT3A activity. In both p.P904-mutated TG01 samples, the amino acid substitution was caused by a CG > TG mutation, raising the possibility that these mutations are related to defective DNA MMR.
Differentially expressed genes are related to proliferation, cell signaling, and developmentTo examine the changes in tumor cell phenotypes and regulation, we performed pairwise differential gene expression analysis with DESeq2 (adjusted p < 0.05, |log2FC| > 1), which resulted in 709 progression-related DE genes (Additional file 1: Table S4). The upregulated genes (n = 386) were linked to the cell cycle, DNA repair, and angiogenesis (ConsensusPathDB and GOrilla gene set enrichment analysis, q < 0.05) (Fig. 1f and Additional file 1: Table S5). Consistently, the progression-related expression signature was dominated by a highly mutually correlated cluster of genes associated with mitotic activity, including the mitotic marker MKI67 (Additional file 2: Fig. S1g). The downregulated genes (n = 323) were associated with cell differentiation, ion transportation, and G protein-coupled receptor (GPCR) signaling (Fig. 1g and Additional file 1: Table S6). Gene set activity analysis (Additional file 1: Table S2) also supported increased proliferation in all progressed grade 4 tumors, except in TG01b (Fig. 1h). Furthermore, hypoxic response, another marker for higher grade, was elevated in all grade 4 tumors in our data (Fig. 1h).
Chromosomal rearrangements are increased after tumor progressionFor chromosomal rearrangement analysis, we called the SVs from the WGS using an in-house tool Breakfast. To separate rearrangements that were more likely to be clonal from the clearly subclonal ones, we further filtered down the SVs based on the fraction of reads supporting the rearrangement. In grade 2–3 astrocytomas, an average of 53 clonal SVs per sample (range 18–87) was identified (Fig. 2a–b). The number of clonal rearrangements was higher in all grade 4 samples than before progression. In cases TG03–TG05, most rearrangements were unique to grade 4 samples (62–79% of clonal SVs in grade 4 samples) whereas TG01 and TG02 grade 4 tumors inherited the majority of clonal SVs (83% and 62%, respectively) from the lower-grade tumor. The pattern was similar for unfiltered SVs, although higher fractions of SVs (42–94%) were unique to grade 4 tumors (Fig. 2a and Additional file 2: Fig. S2a). This suggests that in cases TG03–TG05, the seeding clone of the progressed tumor had obtained novel rearrangements, whereas many cells and their alterations in grade 2–3 tumors were depleted due to surgery and other treatments. Interestingly, the lowest grade 4-unique SV counts, both filtered and unfiltered, were detected in the hypermutator TG01 tumor. Overall, approximately half (41–67% in clonal and 34–74% in all) of the rearrangements were focal (< 2 Mb), affecting proximal regions of the genome, but also interchromosomal translocations were detected (18–38% of clonal and 20–43% of all rearrangements) (Fig. 2b and Additional file 2: Fig. S2a).
Fig. 2
Novel SVs, NRG3 alterations, and decreased NRG3 expression were associated with tumor progression. a The number of SVs was increased in grade 4 tumors. Clonal SVs (upper panel) were used in the subsequent analyses since they are more likely to contribute to progression. b Visualization of clonal SVs with a Circos plot. c In the ICGC cohort, the number of SVs was associated with tumor aggressiveness and recurrence. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001 based on Wilcoxon rank-sum test. d Intragenic rearrangements hit NRG3 gene in two grade 4 tumors. The first exon containing the transcription start site was inverted in TG01b, and a section of the first intron was deleted in TG05b. e NRG3 RNA expression decreased in all the tumors upon progression to grade 4 (p = 0.0022, Wilcoxon rank-sum test). f Representative areas show decreased NRG3 protein expression (red) in IDHmut cells (green) in TG01b. Nuclei are stained with DAPI (blue). g NRG3 level in IDHmut cells was decreased upon progression to grade 4. Quantification of mIHC data across cells in three representative areas per sample. The mean expression for the samples was 1.67, 1.40, and 0.94 for the primary TG01, TG01a, and TG01b tumors, respectively. ****p < 0.0001 based on the Wilcoxon rank-sum test. h NRG3 expression was decreased by tumor grade, especially in grade 4 (p < 0.0001), in 595 TCGA diffuse glioma cases. *p < 0.05, ***p < 0.001, and ****p < 0.0001 based on the Wilcoxon rank-sum test. i Lower NRG3 RNA expression (below 7.7, mean in the entire TCGA diffuse glioma cohort) was associated with shorter OS in TCGA IDHmut astrocytomas (p < 0.0001, log-rank test). j In TCGA IDHmut astrocytoma data, NRG3 was recurrently hemizygously deleted, and its expression was more strongly decreased than PTEN in grade 4 IDHmut astrocytomas. Rearrangements close to genes (*) were observed in cases with gene loss
A similar rearrangement analysis was performed for the SV data from the ICGC cohort including both primary and recurrent diffuse gliomas. Among IDHmut astrocytomas, the highest number of SVs was detected in a recurrent grade 4 tumor with prior radiation, whereas SV counts were similar in primary grade 2 and 4 samples (Fig. 2c). IDHwt GBs had significantly higher clonal SV counts than IDHmut tumors (p = 6.3*10–3 and p = 1.6*10–6 in respect to IDHmut astrocytomas and oligodendrogliomas, respectively, Wilcoxon rank-sum test), and the counts of all SVs were higher in recurrent than primary GBs (p = 0.010, Wilcoxon rank-sum test) (Fig. 2c).
NRG3 rearrangements and decreased expression are associated with tumor progressionChromosomal rearrangements can alter the gene structure or influence gene regulation e.g., by altering regulatory regions or reorganizing TAD structures [31, 32]. TAD boundaries are insulated, and regulatory regions rarely interact with promoters across TAD boundaries [33, 34]. Next, we examined the effects of recurrent chromosomal rearrangements on gene regulation. Fourteen genes were either downregulated (seven genes) or upregulated (seven genes) upon tumor progression, with an accompanying chromosomal rearrangement within the same TAD in at least two cases (Additional file 1: Table S7). For most of these fourteen genes, similar progression-related expression differences were also detected in cases lacking rearrangements (Additional file 1: Table S7). Neuregulin 3 (NRG3) gene, located on chr10q23.1, is of particular interest. Progression-related intragenic NRG3 rearrangements were detected in two cases: a deletion in the first intron in case TG05 and an inversion disrupting the first exon in case TG01 (Fig. 2d); both included putative regulatory regions. Furthermore, TG03b harbored a hemizygous deletion in NRG3. NRG3 was significantly downregulated in all patients upon progression to grade 4 (log2FC = − 3.5, DESeq2 adj. p = 4.2*10–7, Wald test). Among the cases with WGS data (TG01–TG05), the expression was lowest in cases with rearrangements (Fig. 2e). Downregulation of NRG3 protein in TG01 was also detected by mIHC (Fig. 2f–g). In the ICGC WGS data, additional rearrangements in NRG3 (one GB) or in the genomic neighborhood of NRG3 (one grade 2 and one grade 4 IDHmut astrocytoma, one GB) were detected. All these cases also showed a hemizygous loss of NRG3 and low gene expression (Additional file 2: Fig. S2b).
In TCGA diffuse glioma data (n = 595, Additional file 1: Table S8), NRG3 RNA expression decreased with tumor grade in IDHmut astrocytomas and oligodendrogliomas (p < 0.05, Wilcoxon rank-sum test), especially upon progression to grade 4 astrocytomas (p < 0.0001, Wilcoxon rank-sum test) (Fig. 2h). NRG3 expression was the lowest in IDHwt GBs (Fig. 2h). Furthermore, low NRG3 expression (below 7.7 i.e. mean expression in the whole TCGA diffuse glioma cohort) was associated with worse survival in the entire diffuse glioma cohort and within IDHmut astrocytomas (p < 0.0001, log-rank test) (Fig. 2i and Additional file 2: Fig. S2c). The NRG3 expression remained significant (hazard ratio 0.56 for the increase of NRG3 expression by its standard deviation, p = 0.024, Wald test) in a multivariate Cox proportional hazard model including also patient age and tumor grade as variables (Additional file 1: Table S9), or when using NRG3 expression alone (hazard ratio 0.49, p = 0.0026, Wald test). In IDHmut astrocytomas, hemizygous NRG3 losses were significantly more frequent in grade 4 than in grade 2–3 tumors (p = 1.7*10–4, Fisher’s exact test) and were associated with decreased NRG3 expression (Additional file 2: Fig. S2d). PTEN, which is located on the same chromosomal arm as NRG3, was also hemizygously deleted in TG03b and TG05b (Additional file 2: Fig. S2e). In TCGA data, NRG3 was concomitantly deleted with PTEN in 30 out of 34 (88%) IDHmut astrocytomas cases with PTEN loss, and in all (n = 197) IDHwt GBs with PTEN loss (Additional file 1: Table S10). In the GLASS data from matched IDHmut astrocytoma tumors of patients with tumor progression to grade 4 (n = 33, Additional File 1: Table S11), NRG3 was hemizygously deleted in 14 cases upon progression to grade 4, 13 of which were concomitant with a PTEN loss (Additional file 1: Table S12, Additional file 2: Fig. S2f). NRG3 showed a more dramatic decrease in expression than PTEN upon progression to grade 4 in our cases (p = 0.0087, Wilcoxon rank-sum test of log2FCs) (Fig. 2e and Additional file 2: Fig. S2e) and in GLASS data (p = 0.024, Wilcoxon rank-sum test of log2FCs) (Additional file 2: Fig. S2g). Similarly, the downregulation of NRG3 was stronger than for PTEN in TCGA IDHmut astrocytomas with increasing grade (Fig. 2h,j and Additional file 2: Fig. S2h) and upon hemizygous gene deletion (Additional file 2: Fig. S2d). Besides NRG3 and PTEN, other genes in chromosome 10q are often hemizygously lost. In our data, altogether 170 genes with TCGA RNA and CNA data were downregulated (log2FC < − 1) in the lost regions of chr10 in TG03b and/or TG05b. In TCGA IDHmut astrocytomas, 9 of these genes, including NRG3, had significantly (p < 0.05, one-sided Wilcoxon rank-sum test, log2FC < − 1) lower expression in cases with hemizygous loss of gene, compared to neutral copy number (Additional file 1: Table S13). Eight of these genes were also significantly downregulated in grade 4 tumors compared to lower grades (p < 0.05, Wilcoxon rank-sum test, log2FC < − 1) (Additional file 1: Table S13). To conclude these results, NRG3 is one of the few genes that is linked to the progression and loss of chromosome 10q in IDHmut astrocytomas.
Recurrent copy number losses after tumor progressionNext, we analyzed DNA copy number losses and gains at a coarse resolution based on WGS coverage. Progression was accompanied by several novel chromosomal gains and losses in all samples except TG01a, in which only one progression-related deletion was detected (Fig. 3a–b). Before progression to grade 4, the highest CNA counts were observed in patient TG05a (Fig. 3b), who had been treated with radiotherapy seven years prior to surgery (Fig. 1b). Notably, losses were more frequent than gains in all grade 2–3 tumors, and their numbers increased more drastically than the gains in progressed grade 4 tumors (Fig. 3b), consistent with previous results that link increased genomic deletions to radiotherapy [8]. A progression-related loss of an overlapping region was detected in at least three cases on chromosomes 4, 9, 13, 14, 22, and X. Only one region harbored recurrent progression-associated gain: the PDGFRA locus on chromosome 4.
Fig. 3
Progression-related CDKN2A/RB1 alterations were associated with postoperative combination therapy and PDGFRA/MET alterations. a DNA copy number patterns of patients TG01–TG05 in grade 2–3 (left column) and grade 4 (right column) tumors. b Number of copy number losses was increased upon progression to grade 4, especially in cases without a hypermutator phenotype. c Summary of tumor characteristics, patient treatment, and genetic alterations in cases TG01–TG06. Narrow columns represent samples not included in the discovery cohort. d CDKN2A and RB1 were homozygously deleted either through simple rearrangements (TG02b) or complex rearrangement patterns. Rearrangements that were already called in grade 2–3 tumors are visualized in gray. Uncalled rearrangements (dashed lines) were aligned to repeat segments (yellow triangles) in TG05b. e Short microhomologous sequences were detected in rearrangements involved in the reported DNA copy number alterations. f Alterations and treatment information in the targeted sequencing validation cohort consisting of IDHmut astrocytomas, including four discovery cohort cases. Only point mutations, amplifications, and full deletions were analyzed. Narrow columns represent samples with no targeted sequencing data. Relapses after progression to grade 4 are not shown. g Patients who underwent both radiation and chemotherapy after surgery were more likely to develop CDKN2A/RB1 inactivation compared to patients who received radiotherapy alone in the combined discovery and targeted sequencing cohort. Only cases with complete treatment information until progression and confirmed event of inactivation upon progression were included. Inactivating alterations in CDKN2A and RB1 are shown separately but were counted together for Fisher´s exact test. h Rearrangement patterns in PDGFRA and MET genes. The rearrangement at the start of PDGFRA amplification in TG02b is located in the centromere and not called because of satellite repeats (yellow triangles). In TG05, PDGFRA was amplified as an extrachromosomal DNA. PTPRZ1-MET fusion was detected from subclonal rearrangements in TG03b. Uncalled and subclonal rearrangements (dashed arrows and arcs) are only visualized when affecting the alteration
In ICGC data, IDHwt GBs had a higher proportion of copy number losses than other tumor types (p = 0.012 and p = 3.4*10–5 with respect to IDHmut astrocytomas and oligodendrogliomas, respectively, Wilcoxon rank-sum test). Losses tended to be more frequent than gains in recurrent tumor samples, most of which were taken post-radiation (p = 0.14, Wilcoxon rank-sum test) (Additional file 2: Fig. S3a). In contrast, gains were more frequent than losses in primary diffuse glioma tumors (p = 0.035, Wilcoxon rank-sum test) (Additional file 2: Fig. S3a).
Inactivation of CDKN2A or RB1 in progressed grade 4 tumors associated with the combination of chemo- and radiotherapyRadiation has been reported to increase the risk of CDKN2A deletion [8]. Furthermore, homozygous CDKN2A deletion has been linked to the recurrence and progression of IDHmut astrocytomas and is currently a determining criterion for a grade 4 diagnosis, irrespective of low-grade tissue histology [3, 10, 35]. Although the original tumor grading was not based on the CDKN2A status in our discovery cohort, CDKN2A was homozygously deleted and dramatically downregulated in four progressed grade 4 tumors (Fig. 3c and Additional file 2: Fig. S3b). Furthermore, a subclonal truncating mutation (frameshift insertion, chr13:48,307,353:- > AG, p.R71fs, AF 0.20), together with a hemizygous loss of RB1, was detected in TG01b and a homozygous deletion of RB1 in TG04b. In summary, either CDKN2A or RB1 was fully deleted upon progression in all cases, except for the hypermutator TG01 with a subclonal RB1 inactivation. This makes inactivation of the CDKN2A/RB1 pathway a major mechanism for grade 4 tumor development, at least with the treatment schemes used.
To study the origin of CNAs, we analyzed the rearrangement patterns that caused CDKN2A and RB1 deletions. TG02b had already lost one copy of CDKN2A in the first analyzed sample and the second copy was deleted via a simple deletion involving two breakpoints (Fig. 3d). TG03b lost the first copy due to a translocation with chromosome X, and the second CDKN2A deletion was related to a fusion between the start of chr9 and an inverted segment (chr9p21.3-chr9q22.33) on the right side of CDKN2A (Fig. 3d). In TG05b, the rearrangement pattern was rather complex, involving translocations with chr12 and the end of chr9, thus linking the CDKN2A deletion to a large gain in chr9q (Fig. 3d). The rearrangements in TG05b were partly connected to the SINE element AluSx across chromosome 9 and were not reported by the detection tool. Similarly, in TG04b, one copy of RB1 was lost in a larger deletion, and the second loss was linked to a very complex rearrangement pattern; 24 SVs were detected in the genomic neighborhood and within the depleted region, suggesting that small stretches of DNA were fused with the regions adjacent to the homozygously deleted region. In cases TG03b–TG05b, short (2–4 bp) microhomologous sequences were detected in a fraction of breakpoints (Fig. 3e) and were indicative of MMEJ or NHEJ pathway activity [36]. In one RB1 rearrangement, the microhomology was 10 bp and was evidently repaired with MMEJ (the sole repair mechanism for breakpoints with 5–18 bp microhomology) [37]. As longer homologies were not detected, HRR was unlikely to be involved.
Targeted DNA sequencing was used to analyze FFPE or fresh frozen tumor material from 21 additional patients with IDHmut astrocytoma who had progressed to grade 4 (Additional file 1: Tables S14-S15). CDKN2A was inactivated in the grade 4 tumors of nine (43%) additional patients; it was homozygously deleted in eight cases and carried a truncating mutation together with hemizygous loss in TG26 (Fig. 3f). No additional RB1 mutations were detected in the targeted sequencing cohort. When considering the inactivation of CDKN2A/RB1 with respect to treatment, patients who underwent both postoperative radiation and chemotherapy were more likely to develop inactivation after therapy than those who underwent only postoperative radiotherapy (p = 0.02, Fisher's exact test) (Fig. 3g). This difference seems unrelated to primary tumor grade (five/six grade 2/3 tumors developed a CDKN2A/RB1 alteration upon progression to grade 4 vs. six/six grade 2/3 tumors without progression-related alteration, respectively, p = 1.0, Fisher’s exact test). Additionally, there were no significant differences in the distribution of grade 2 and 3 tumors regarding the given treatment in this cohort. Only one (TG07) of the seven cases with CDKN2A/RB1 inactivation after combination therapy received TMZ before the gene inactivation. TP53 is typically defective in IDHmut astrocytomas, which was also true for all but four cases in our discovery and targeted sequencing cohorts (Fig. 3c, f). TP53 alterations are likely to disturb the TP53-dependent cell cycle checkpoint pathway, induced by DNA damage [38]. In all cases with intact TP53 in our in-house cohorts, progression to grade 4 disease was accompanied by CDKN2A inactivation.
As expected, recurrent homozygous deletions in CDKN2A and RB1 were present in TCGA diffuse gliomas and GLASS progressed grade 4 IDHmut astrocytomas (Additional file 1: Table S8 and Table S11 and Additional file 2: Fig. S3c and S2f). In GLASS data, CDKN2A/RB1 inactivation was the most frequent progression-related alteration, with 18 out of 33 analyzed cases having full inactivation of CDKN2A, two cases having fully inactivated RB1, and one case with a truncating mutation in one RB1 allele (Additional file 1: Table S12, Additional file 2: Fig. S2f). Subfraction of these gene inactivations were caused by the combined effect of hemizygous loss and a damaging point/frameshift mutation in the other allele. All the damaging point mutations represented the TMZ-associated mutational signature [27] and were detected after TMZ therapy. Both within GLASS and TCGA IDHmut astrocytomas, the inactivation of CDKN2A and RB1 was mainly mutually exclusive. In TCGA, one grade 4 tumor had both CDKN2A and RB1 called as homozygously deleted but only RB1 showed decreased expression (Additional file 2: Fig. S3c). In GLASS, for one case with a similar alteration pattern in these genes no expression data was available. In TCGA, the majority of grade 4 IDHmut astrocytomas (68%) had a homozygous deletion of CDKN2A, which also led to reduced expression at the RNA level. RB1 was homozygously deleted in three grade 2–3 IDHmut astrocytomas with poor OS rates (Additional file 2: Fig. S3c). CDKN2A or RB1 expression was not associated with survival in diffuse glioma subtypes (Additional file 2: Fig. S3d–e).
Progression-related activating alterations in growth factor receptor genes concomitant with CDKN2A inactivationGrade 4 tumors showed mutually exclusive activating alterations in the growth factor receptor genes (Fig. 3c). PDGFRA was amplified in TG02b (14 extra copies) and TG05b (50 extra copies). In TG02b, amplification started at a satellite repeat region in the centromere (Fig. 3h). In TG05b, the amplified region included a short 9 kb stretch in chr4p16.1, forming an extrachromosomal DNA amplicon (Fig. 3h). Subclonal PDGFRA amplification with identical breakpoints was also detected in TG05a (taken post-radiotherapy), suggesting that this subclone was the seeding clone for the grade 4 tumor. In our targeted sequencing cohort, PDGFRA amplification was observed in four additional cases (TG07, TG14, TG18, and TG25) (Fig. 3f). In TG18, amplification was already present in the primary grade 3 tumor and was not coupled with CDKN2A deletion; in the others, amplification was acquired upon progression, concomitantly with CDKN2A deletion. All these four cases received radiation and two received additional chemotherapy before progression to grade 4. Furthermore, TG03b acquired a subclonal MET-PTPRZ1 fusion (Fig. 3h). A striking overexpression of the gene was detected in discovery cohort cases with activating alterations in PDGFRA or MET (Additional file 2: Fig. S3f).
In TCGA IDHmut astrocytomas, PDGFRA amplification occurred more often in grade 4 tumors (21% in grade 4, 0.03% in grade 2–3, p = 0.0025, Fisher’s exact test) (Additional file 2: Fig. S3c). Even when considering generally higher PDGFRA expression in IDHmut astrocytomas than in IDHwt GBs (Additional file 2: Fig. S3c), only a subpopulation of tumors with gene amplification represented outlier expression levels (in IDHmut astrocytomas, four tumors with PDGFRA amplification, three of which were grade 4).
Of the 21 additional patients in the targeted sequencing cohort, 11 (52%) showed no CDKN2A, RB1, or PDGFRA alterations. Eight (73%) of these, including TG22 with a truncating mutation in RAD52, MSH2, and MSH6 DNA repair genes, received radiation alone before progression to grade 4 (Fig. 3f). Activating PDGFRA/MET alterations always co-occurred with a homozygous CDKN2A deletion in our discovery cohort and in all but one case in the targeted sequencing cohort (only PDGFRA amplification in TG18) (p = 0.033 for concomitant PDGFRA/CDKN2A alteration in the combined discovery and targeted seq cohort, Fisher’s exact test). Furthermore, CDKN2A was completely inactivated in all the grade 4 IDHmut astrocytomas with PDGFRA amplification in TCGA and GLASS data with an exception of two cases (one in the GLASS and one in TCGA dataset), which carried only a PDGFRA amplification (Additional file 2: Fig. S2f and S3c). However, these two samples also showed no increase in the PDGFRA expression (Additional file
Comments (0)