
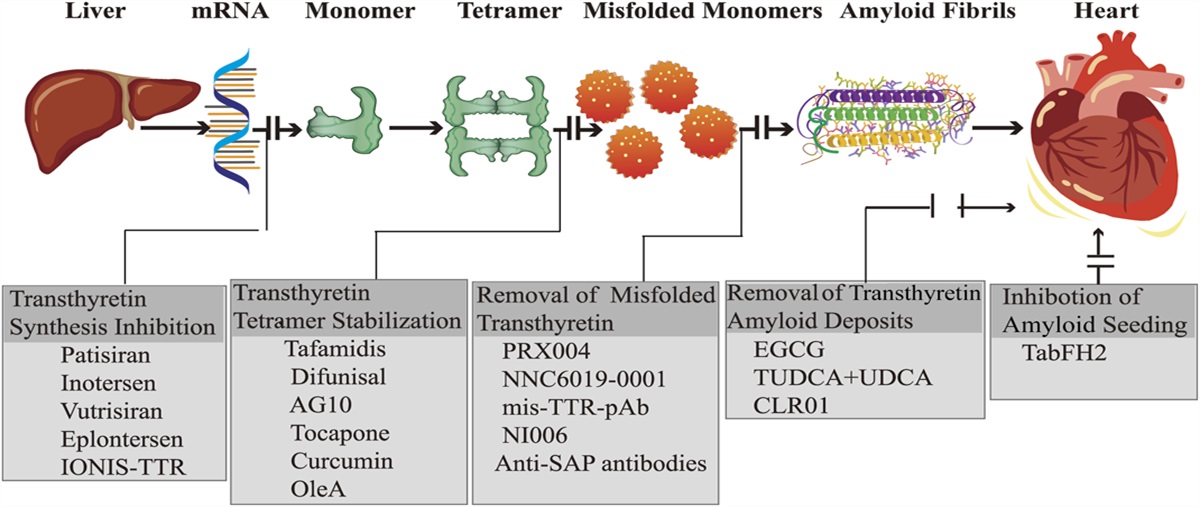
記住我

Visfatin/nicotinamide phosphoribosyltransferase (NAMPT) was first discovered as a cytokine named pre-B-cell colony-enhancing factor and later rediscovered as an important enzyme in nicotinamide adenine dinucleotide generation, which has considerably widened its potential biological activities.1 Although visfatin/NAMPT was originally thought to be produced exclusively by the adipose tissue (adipocytes and infiltrating macrophages), other tissues such as skeletal muscle, liver, and brain as well as cardiomyocytes and immune cells are also able to synthetize it.1 Noteworthy, both the cytokine-like (extracellular) and enzymatic (intracellular) functions seem to be responsible for its relevance in immunity, metabolism, and stress responses in both physiological and pathological conditions.1 Several studies have shown that the extracellular NAMPT (eNAMPT) participates in various cellular functions such as apoptosis, inflammation, and extracellular matrix degradation; however, there is debate regarding the underlying mechanisms for these effects, as reviewed elsewhere.1
Growing evidence indicates that the circulating levels of visfatin are altered in cardiovascular diseases, including atherosclerosis, essential hypertension, and preeclampsia.2 Curiously, the association between visfatin levels with circulating markers of endothelial dysfunction and with systolic and diastolic blood pressure levels seems to be independent of the degree of adiposity.3,4 However, the role of visfatin in mediating vascular dysfunction and hypertension are still controversial.
In this context, Bayram et al have recently published a study aiming to investigate the effects of visfatin on vascular responses in the human left internal mammary artery, the gold standard vessel for coronary artery bypass grafting, with the possible underlying mechanisms.5 They reported that visfatin did not significantly affect the contractile responses of internal mammary artery rings to several vasoconstrictors. However, visfatin induced relaxation of internal mammary artery rings preconstricted by phenylephrine in an endothelium-dependent manner, and this vasodilatory response was attenuated by incubation with a selective guanylate cyclase inhibitor and with a NAMPT enzymatic inhibitor FK866.5 Although we agree that these findings may have a substantial importance for new therapeutic approaches toward coronary artery disease, we would like to further discuss the results regarding the effects of visfatin on endothelium-dependent vasorelaxation. We do not agree with the authors when they interpreted that incubation of internal mammary artery rings preconstricted by phenylephrine with visfatin in the 10−9 and 10−8 M concentrations for 30 minutes decreased the relaxant responses to cumulatively applied concentrations of acetylcholine. Figure 6 and Table 1 in this study from Bayram et al5 actually show that the control and visfatin at 10−10 M groups relaxed around 70% in response to acetylcholine, and the incubation with visfatin at 10−9 and 10−8 M increased acetylcholine-induced maximal relaxation to around 90%. These observations are in line with a prior study demonstrating a vasodilatory effect of visfatin (100 ng/mL) in noradrenaline-preconstricted aorta of male Wistar rats through endothelium-derived nitric oxide (NO).6
Conversely, several studies point to visfatin as a factor associated with endothelial dysfunction. Indeed, the same research group has previously performed a similar set of experiments but using small mesenteric arteries isolated from male Wistar rats instead.7 They found that visfatin incubation (at doses ranging from 1 to 100 ng/mL) reduced the relaxation responses to acetylcholine in phenylephrine-preconstricted endothelium-intact mesenteric rings, and this effect was reverted by the incubation with NAMPT enzymatic inhibitor FK866 or superoxide dismutase.7 In addition, visfatin was shown to impair endothelium-dependent vasodilation in these vessels through a reduction in NO bioavailability caused by increased formation of superoxide anions.7 Similar results were observed in bovine small coronary arteries precontracted with phenylephrine.8 Moreover, visfatin is also able to decrease endothelium-dependent vasorelaxation partially by nicotinamide adenine dinucleotide phosphate hydrogen oxidase stimulation in rat (male Sprague–Dawley) and human (female) mesenteric vessels.9 Further evidence in humans (male) for the detrimental actions of visfatin in the endothelium is that obese individuals present lower quantity of endothelial progenitor cells along with higher visfatin and oxidative stress products in the circulation than controls.10 Taken together, these reports suggest that nicotinamide adenine dinucleotide phosphate hydrogen oxidase–induced release of superoxide anions may play a role in visfatin-induced impaired NO-mediated vasodilation, especially under obesity conditions.
Nonetheless, in agreement with a possible dilatory effect of visfatin on the vasculature, Zhou et al11 have recently assessed visfatin levels in aorta of patients undergoing aortic replacement surgery due to acute aortic dissection. In this study, NAMPT expression was found to be reduced in the aortic tissue of patients with hypertension compared with those with normal blood pressure levels. Moreover, male NAMPT-knockout mice exhibited higher blood pressure and reactive oxygen species levels after continuous infusion of angiotensin II for 4 weeks compared with wild-type counterparts, and administration of recombinant human NAMPT prevented these effects of angiotensin II on blood pressure and reactive oxygen species levels. Treatment with recombinant human NAMPT also improved angiotensin II–induced abnormal vascular remodeling in NAMPT knockout mice, suggesting NAMPT as a potential therapeutic target for hypertension.
However, another recent in vivo study stands against these findings by showing that small mesenteric arteries of male C57BL/6 mice continuously infused with visfatin/eNAMPT for 7 days (100 ng/kg per day) presented reduced endothelium-dependent relaxation in noradrenaline precontracted rings.12 The coinfusion of visfatin/eNAMPT with NAMPT inhibitor FK866 or toll-like receptor (TLR)-4 blocker CLI095 attenuated this vasoconstrictor effect, supporting a role for NAMPT enzymatic activity and TLR4 activation in endothelial dysfunction induced by visfatin/eNAMPT. Furthermore, visfatin/eNAMPT-induced endothelial dysfunction was improved by the nod-like receptor protein 3 (NLRP3)–inflammasome inhibitor MCC950 and by the interleukin (IL)-1-receptor antagonist anakinra, suggesting that IL-1β released by NLRP3-inflammasome may also mediate endothelial damage caused by visfatin/eNAMPT.
Interestingly, the potential interaction among eNAMPT, TLR-4, and cytokines, including NLRP3 in the pathophysiology of preeclampsia, has been reviewed elsewhere.13 Thus, we would like to close this commentary by discussing the potential role of visfatin endothelial dysfunction associated with preeclampsia. Placental ischemia/hypoxia leads to the release of antiangiogenic factors into maternal circulation, including the soluble fms-like tyrosine kinase (sFLT)-1, which is an important mediator of widespread maternal endothelial dysfunction in preeclampsia by blocking the effects of vascular endothelial growth factor on angiogenesis and vascular health.14 Moreover, preeclampsia is characterized by decreased circulating levels of NO, which was shown to be inversely related to sFLT‐1.15 Although most studies provide evidence for increased circulating levels of visfatin in preeclampsia, there are few reports of similar or even decreased visfatin levels in preeclampsia as compared with normotensive pregnant women.16 Nevertheless, visfatin/NAMPT levels were shown to be inversely related to NO but positively related to sFLT-1 levels in the circulation of pregnant women with preeclampsia.17 Similar relationships between visfatin/NAMPT levels with NO and soluble sFLT-1 were observed in the subgroup of patients with preeclampsia classified as nonresponsive to antihypertensive therapy, who exhibited higher levels of sFLT-1 and the worst clinical outcomes as compared with the subgroup of patients with preeclampsia classified as responsive to antihypertensive therapy.18 Although these findings may suggest that visfatin/NAMPT inhibits NO formation and upregulates the production of sFLT-1 in preeclampsia, the potential contribution of visfatin/NAMPT to endothelial dysfunction in this hypertensive disorder of pregnancy needs to be confirmed by additional studies.
Other relevant variables related to experimental design that should be considered when evaluating results of vasocontractility studies is concentration of substances/drugs because they frequently use concentrations that are different from those found/used in human diseases, and exposure time, as vessels are incubated with these substances/drugs acutely (usually 1–2 hours), which also differs from the chronic exposure that vessels undergo during the course of chronic syndromes (months to years). Ex vivo studies are important to guide the vasoactive role of a substance but do not exactly reflect what happens in vivo most of the time.
In conclusion, although the mechanisms underlying the possible beneficial and detrimental actions of visfatin on the vasculature are still being uncovered, visfatin definitely plays a role in inflammation and endothelial function (Fig. 1). It seems that the contribution of visfatin to the development or attenuation of pathological processes implicated in coronary artery disease and hypertension depends on its concentration (low vs. high), exposure time (acute vs. chronic), and the vascular bed (resistant vs. complaint, healthy vs. sick) being examined; however, these questions still need to be addressed by further studies.
 FIGURE 1.: Vascular mechanisms underlying the effects of visfatin/eNAMPT on endothelium in normal vascular homeostasis (blue shaded area), and the potential detrimental actions of visfatin/eNAMPT on vascular function (red shaded area), such as stimulation of NADPH oxidase,9 exaggerated ROS levels,11 reduction in NO bioavailability caused by oxidative stress,7 and activation of TLR-4 with downstream formation of NLRP3 inflammasome.12 These visfatin/eNAMPT-stimulated mechanisms may contribute to amelioration or development of pathological processes implicated in coronary artery disease and hypertension. Dotted arrows were used to illustrate the potential visfatin/eNAMPT stimulation of sFLT-1 synthesis, with subsequent reductions in VEGF and NO levels; however, this pathway of visfatin/eNAMPT on mediating vascular dysfunction and hypertension during pregnancy needs to be confirmed by further studies. NADPH, nicotinamide adenine dinucleotide phosphate hydrogen; ROS, reactive oxygen species; VEGF, vascular endothelial growth factor.
1. Dahl TB, Holm S, Aukrust P, et al. Visfatin/NAMPT: a multifaceted molecule with diverse roles in physiology and pathophysiology. Annu Rev Nutr. 2012;32:229–243.
2. Filippatos TD, Randeva HS, Derdemezis CS, et al. Visfatin/PBEF and atherosclerosis-related diseases. Curr Vasc Pharmacol. 2010;8:12–28.
3. Gunes F, Akbal E, Cakir E, et al. Visfatin may be a novel marker for identifying stages of essential hypertension in advanced age patients. Intern Med. 2012;51:553–557.
4. Reimann M, Ziemssen T, Huisman HW, et al. Ethnic-specific correlations of visfatin with circulating markers of endothelial inflammation and function. Obesity (Silver Spring). 2009;17:2210–2215.
5. Bayram Z, Akcabag E, Ozbey G, et al. The functional effects of visfatin on human left internal mammary artery. J Cardiovasc Pharmacol. 2022;80:725–731.
6. Yamawaki H, Hara N, Okada M, et al. Visfatin causes endothelium-dependent relaxation in isolated blood vessels. Biochem Biophysical Res Commun. 2009;383:503–508.
7. Akcabag E, Bayram Z, Kucukcetin IO, et al. Functional effects of visfatin in isolated rat mesenteric small resistance arteries. Eur J Pharmacol. 2021;908:174333.
8. Xia M, Zhang C, Boini KM, et al. Membrane raft-lysosome redox signalling platforms in coronary endothelial dysfunction induced by adipokine visfatin. Cardiovasc Res. 2011;89:401–409.
9. Vallejo S, Romacho T, Angulo J, et al. Visfatin impairs endothelium-dependent relaxation in rat and human mesenteric microvessels through nicotinamide phosphoribosyltransferase activity. PLoS One. 2011;6:e27299.
10. Chen S, Sun L, Gao H, et al. Visfatin and oxidative stress influence endothelial progenitor cells in obese populations. Endocr Res. 2015;40:83–87.
11. Zhou L, Zhang S, Bolor-Erdene E, et al. NAMPT/SIRT1 attenuate Ang II-induced vascular remodeling and vulnerability to hypertension by inhibiting the ROS/MAPK pathway. Oxid Med Cell Longev. 2020;2020:1–21.
12. Romacho T, Valencia I, Ramos-Gonzalez M, et al. Visfatin/eNAMPT induces endothelial dysfunction in vivo: a role for Toll-Like Receptor 4 and NLRP3 inflammasome. Sci Rep. 2020;10:5386.
13. Nunes PR, Ceron CS, Luizon MR, et al. Interaction among extracellular nicotinamide phosphoribosyltransferase, toll-like receptor-4, and inflammatory cytokines in pre-eclampsia. Am J Reprod Immunol. 2022;87:e13514.
14. Ceron CS, Pereira DA, Sandrim VC, et al. Potential roles of visfatin/NAMPT on endothelial dysfunction in preeclampsia and pathways underlying cardiac and vascular remodeling. J Cell Physiol. 2022;237:10–12.
15. Sandrim VC, Palei AC, Metzger IF, et al. Nitric oxide formation is inversely related to serum levels of antiangiogenic factors soluble fms-like tyrosine kinase-1 and soluble endogline in preeclampsia. Hypertension. 2008;52:402–407.
16. Luizon MR, Belo VA, Palei AC, et al. Effects of NAMPT polymorphisms and haplotypes on circulating visfatin/NAMPT levels in hypertensive disorders of pregnancy. Hypertens Res. 2015;38:361–366.
17. Pereira DA, Sandrim VC, Palei ACT, et al. NAMPT levels are inversely related to nitric oxide formation and positively related to soluble fms-like tyrosine kinase-1 levels in preeclampsia. Pregnancy Hypertens. 2019;18:137–140.
18. Pereira DA, Sandrim VC, Palei AC, et al. NAMPT single-nucleotide polymorphism rs1319501 and visfatin/NAMPT affect nitric oxide formation, sFlt-1 and antihypertensive therapy response in preeclampsia. Pharmacogenomics. 2021;22:451–464.
FIGURE 1.: Vascular mechanisms underlying the effects of visfatin/eNAMPT on endothelium in normal vascular homeostasis (blue shaded area), and the potential detrimental actions of visfatin/eNAMPT on vascular function (red shaded area), such as stimulation of NADPH oxidase,9 exaggerated ROS levels,11 reduction in NO bioavailability caused by oxidative stress,7 and activation of TLR-4 with downstream formation of NLRP3 inflammasome.12 These visfatin/eNAMPT-stimulated mechanisms may contribute to amelioration or development of pathological processes implicated in coronary artery disease and hypertension. Dotted arrows were used to illustrate the potential visfatin/eNAMPT stimulation of sFLT-1 synthesis, with subsequent reductions in VEGF and NO levels; however, this pathway of visfatin/eNAMPT on mediating vascular dysfunction and hypertension during pregnancy needs to be confirmed by further studies. NADPH, nicotinamide adenine dinucleotide phosphate hydrogen; ROS, reactive oxygen species; VEGF, vascular endothelial growth factor.
1. Dahl TB, Holm S, Aukrust P, et al. Visfatin/NAMPT: a multifaceted molecule with diverse roles in physiology and pathophysiology. Annu Rev Nutr. 2012;32:229–243.
2. Filippatos TD, Randeva HS, Derdemezis CS, et al. Visfatin/PBEF and atherosclerosis-related diseases. Curr Vasc Pharmacol. 2010;8:12–28.
3. Gunes F, Akbal E, Cakir E, et al. Visfatin may be a novel marker for identifying stages of essential hypertension in advanced age patients. Intern Med. 2012;51:553–557.
4. Reimann M, Ziemssen T, Huisman HW, et al. Ethnic-specific correlations of visfatin with circulating markers of endothelial inflammation and function. Obesity (Silver Spring). 2009;17:2210–2215.
5. Bayram Z, Akcabag E, Ozbey G, et al. The functional effects of visfatin on human left internal mammary artery. J Cardiovasc Pharmacol. 2022;80:725–731.
6. Yamawaki H, Hara N, Okada M, et al. Visfatin causes endothelium-dependent relaxation in isolated blood vessels. Biochem Biophysical Res Commun. 2009;383:503–508.
7. Akcabag E, Bayram Z, Kucukcetin IO, et al. Functional effects of visfatin in isolated rat mesenteric small resistance arteries. Eur J Pharmacol. 2021;908:174333.
8. Xia M, Zhang C, Boini KM, et al. Membrane raft-lysosome redox signalling platforms in coronary endothelial dysfunction induced by adipokine visfatin. Cardiovasc Res. 2011;89:401–409.
9. Vallejo S, Romacho T, Angulo J, et al. Visfatin impairs endothelium-dependent relaxation in rat and human mesenteric microvessels through nicotinamide phosphoribosyltransferase activity. PLoS One. 2011;6:e27299.
10. Chen S, Sun L, Gao H, et al. Visfatin and oxidative stress influence endothelial progenitor cells in obese populations. Endocr Res. 2015;40:83–87.
11. Zhou L, Zhang S, Bolor-Erdene E, et al. NAMPT/SIRT1 attenuate Ang II-induced vascular remodeling and vulnerability to hypertension by inhibiting the ROS/MAPK pathway. Oxid Med Cell Longev. 2020;2020:1–21.
12. Romacho T, Valencia I, Ramos-Gonzalez M, et al. Visfatin/eNAMPT induces endothelial dysfunction in vivo: a role for Toll-Like Receptor 4 and NLRP3 inflammasome. Sci Rep. 2020;10:5386.
13. Nunes PR, Ceron CS, Luizon MR, et al. Interaction among extracellular nicotinamide phosphoribosyltransferase, toll-like receptor-4, and inflammatory cytokines in pre-eclampsia. Am J Reprod Immunol. 2022;87:e13514.
14. Ceron CS, Pereira DA, Sandrim VC, et al. Potential roles of visfatin/NAMPT on endothelial dysfunction in preeclampsia and pathways underlying cardiac and vascular remodeling. J Cell Physiol. 2022;237:10–12.
15. Sandrim VC, Palei AC, Metzger IF, et al. Nitric oxide formation is inversely related to serum levels of antiangiogenic factors soluble fms-like tyrosine kinase-1 and soluble endogline in preeclampsia. Hypertension. 2008;52:402–407.
16. Luizon MR, Belo VA, Palei AC, et al. Effects of NAMPT polymorphisms and haplotypes on circulating visfatin/NAMPT levels in hypertensive disorders of pregnancy. Hypertens Res. 2015;38:361–366.
17. Pereira DA, Sandrim VC, Palei ACT, et al. NAMPT levels are inversely related to nitric oxide formation and positively related to soluble fms-like tyrosine kinase-1 levels in preeclampsia. Pregnancy Hypertens. 2019;18:137–140.
18. Pereira DA, Sandrim VC, Palei AC, et al. NAMPT single-nucleotide polymorphism rs1319501 and visfatin/NAMPT affect nitric oxide formation, sFlt-1 and antihypertensive therapy response in preeclampsia. Pharmacogenomics. 2021;22:451–464.
留言 (0)