
Remember me



Patient 1 (P1), a 20-year-old Jewish male, was treated at the Allergy and Clinical Immunology Unit at Hadassah Medical Center, Jerusalem, Israel. He was born to non-consanguineous, healthy parents and is the only symptomatic child among five healthy siblings (Fig. 1A). Physical examination revealed dysmorphic features, including hypertelorism, microcephaly, a broad nasal bridge, and coarse facial features.
Fig. 1
Clinical manifestations of the patient. (A) The family pedigree of patient (P)1 demonstrating a de-novo variant in a non-consanguineous family with 4 unaffected siblings. (B-D) skin biopsy pathology of P1; Punch of skin with flattened epidermis with compact hyper keratosis (B). In dermis- fibrosis, mild chronic inflammation including plasma cells (C). In the background- few eosinophils (D). The findings are consistent with chronic atopic dermatitis (H&E X 400). (E, F) Platelet count and mean platelet volume of P1 overtime demonstrating persistent macro-thrombocytopenia. (G, H) Severe purulent impetigo, as seen in P1’s lower limbs and scalp, respectively
The patient has chronic allergic rhinitis and severe atopic dermatitis with prurigo nodularis, which are treated with topical steroids and phototherapy (Fig. 1B-D). He also has chronic diarrhea that has not been fully evaluated or treated. His medical history includes intellectual impairment, failure to thrive, and developmental delay. Additionally, he was diagnosed with short stature characterized by a bone age delay of two years. His growth hormone levels following glucagon and arginine stimulation tests were within the normal range. Interestingly, his blood tests from early childhood revealed consistent thrombocytopenia with unusually large platelets (see Fig. 1E for platelet counts and Fig. 1 F for mean platelet volumes).
P1’s medical history is also notable for recurrent infections. He had recurrent pneumococcal pneumonia and bacteremia, including two admissions to the pediatric intensive care unit at ages 4 and 5. He also had recurrent episodes of viral acute otitis media, streptococcal periorbital cellulitis, and severe purulent impetigo (Fig. 1G, H).
His pulmonary workup included a chest computed tomography scan, which demonstrated emphysematous changes in both lung apices; diffuse thickening of air bronchi, and atelectasis of segments in the right middle and lower lobes. Spirometry revealed an obstructive disorder with a forced expiratory volume (FEV1) to forced volume capacity (FVC) ratio of 0.75. An increase in FEV1 by 14% following salbutamol treatment confirmed the diagnosis of mild asthma.
Overall, P1’s complex medical history underscores the need for a thorough evaluation and targeted management of significant immunological and developmental challenges.
Baraitser-Winter Syndrome Type 1 in the Patient is Caused by a Novel ACTB VariantNext, using exome sequencing we identified a cytidine duplication at position 1078 in exon 6 of the ACTB gene, resulting in a novel heterozygous variant, c.1078dupC, p.Gln360ProfsTer4 (Fig. 2 A). This duplication leads to a frameshift that introduces a premature stop codon downstream (Ter 4), resulting in a truncated β-actin protein consisting of 362 amino acids, compared to the full-length protein of 375 amino acids (Fig. 2B). The p.Gln360ProfsTer4 variant is novel and absent from local or global databases (seenB4 score of 0). It is classified as likely pathogenic by different prediction softwares using the American College of Medical Genetics (ACMG) guidelines. Family segregation studies via Sanger sequencing revealed that neither parent of P1 harbored the ACTB variant, therefore characterizing it as de-novo. A cryo-electron microscopy model of β-actin in the patient and a healthy control (HC) is presented in Fig. 2C. Analysis of the truncated β-actin reveals alterations in the C-terminal domain, a critical component in actin filament dynamics and protein-protein interactions. These structural changes in the truncated β-actin may disrupt actin polymerization, stability, or binding to regulatory proteins, thus indicating possible pathogenic characteristics of the ACTB variant.
Fig. 2
ACTB variant found in the patient. (A) Sanger sequencing of P1 and a healthy control demonstrates a heterozygous c.1078dupC, p.Gln360ProfsTer4 nonsense variant in Exon 6 of ACTB. (B) Position of the variant leading to premature stop codon and truncated β-actin protein (C) The cryo- electron microscopy model of β-actin in the patient and a HC was compiled using the PyMOL Molecular Graphics System, Version 3.0, Schrödinger, LLC. (protein data bank: 5JLH [49])
The p.Gln360ProfsTer4 β-Actin Subject to Enhanced Cellular DegradationWe then sought to evaluate the pathogenicity of the p.Gln360ProfsTer4 ACTB variant by comparing the overall β-actin protein expression levels in protein extracts of peripheral blood mononuclear cells (PBMCs) from P1 and HC using immunoblotting. This analysis revealed a marked decreased β-actin expression in P1 compared to HCs (Fig. 3 A,B).
Fig. 3
The p.Gln360ProfsTer4 β-actin protein expression and stability. (A, B) PBMC were lysed, protein extracts were separated by SDS-PAGE, transferred into a nitrocellulose membrane and tested for β-actin. GAPDH was used as a loading control. Immunoblotting has revealed that although GAPDH was increased in P1 vs. HC, expression of β-actin was nearly absent in the patient. (C, D) Human embryonic kidney 293 T (HEK293T) cells were transfected with plasmids encoding HA-β-actin (WT) and FLAG-β-actin (p.Gln360ProfsTer4 variant). Forty-eight hours post-transfection, cells were treated with cycloheximide and harvested at 0, 6, 12, 24, 30, and 48 h following treatment. The levels of exogenous WT and mutant β-actin were assessed for each time point by immunoblotting
The p.Gln360ProfsTer4 variant likely affects residues 360 and beyond, including regions involve in actin polymerization (Fig. 3B) [12]. Additionally, the anti-β-actin antibody used for immunoblotting targets the same N-terminal amino acid sequence in β-actin proteins from both HC and P1 [13]. Therefore, the decrease in the β-actin band in P1’s immunoblotting may suggest enhanced degradation of the mutant β-actin. To test this, we compared the half-lives of the mutant and WT β-actin. HEK293T cells were transfected with expression plasmids for HA-β-actin (WT) and FLAG-β-actin (p.Gln360ProfsTer4 variant). Forty-eight hours post-transfection, the cells were treated with the potent eukaryotic translation elongation inhibitor cycloheximide (CHX) [14] and harvested at 0, 6, 12, 24, 30, and 48 h after treatment. We then determined by immunoblotting the levels of exogenous WT or mutant β-actin, as well as total β-actin, using antibodies against FLAG, HA, and β-actin, respectively. The expression of mutant β-actin was diminished by six hours post-CHX treatment, while the levels of WT β-actin and endogenous β-actin remained stable for up to 48 h (Fig. 3C, D).
These results indicate that the half-life of the p.Gln360ProfsTer4 variant is significantly shorter than that of WT β-actin, which may explain the reduced expression observed in PBMCs from P1 (Fig. 4A, B).
Fig. 4
Impaired cytoskeletal localization of mutant β-actin (p.Gln360ProfsTer4) and its dominant-negative effect. (A) Human embryonic kidney 293 T (HEK293T) cells were transfected with expression plasmids encoding HA-tagged wild-type (WT) β-actin, FLAG-tagged mutant β-actin (p.Gln360ProfsTer4), or both. The cells were than processed to complete immunoblotting for transfection conformation and confocal microscopy of HA and FLAG. (B) Immunoblotting of HA and FLAG demonstrating successful transfection of WT plasmid, mutant plasmid and both. (C-D) Confocal microscopy analysis of actin cytoskeleton organization. Cells were stained with DAPI (nuclei, blue), phalloidin-iFluor 488 (endogenous β-actin, green), and antibodies against HA (WT exogenous β-actin, red) and FLAG (mutant exogenous β-actin, magenta). Periphery-to-whole-cell fluorescence standard deviation distribution was quantified in FLAG- and HA-tagged cells, representing exogenous actin distribution in WT and mutant cells. (E) Quantification of co-localization between exogenous and endogenous β-actin. (F-G) Periphery-to-whole-cell fluorescence standard deviation ratio and co-localization with endogenous β-actin in HEK293T cells co-transfected of WT and mutant β-actin. Each dot represents an individual cell. The two-tailed, unpaired Mann–Whitney test was used for statistical analysis. Results are presented as mean ± SD (P values: ns = non-significant; *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001)
Validating the Dominant-Negative Effect of the ACTB Variant via In-Vitro Transfection ModelTo further investigate the pathogenicity of the p.Gln360ProfsTer4 β-actin variant, we examined its effect on the actin cytoskeleton. HEK293T cells were transfected with expression plasmids for HA-tagged WT β-actin, FLAG-tagged mutant β-actin (p.Gln360ProfsTer4), or both (Fig. 4 A). Successful transfection was confirmed by immunoblotting (Fig. 4B). Confocal microscopy was used to analyze the distribution of exogenous and endogenous β-actin. Cells were stained with DAPI (nuclei), phalloidin-iFluor 488 (endogenous β-actin), and antibodies against HA and FLAG (exogenous β-actin). Transfection with the FLAG-tagged mutant β-actin plasmid resulted in a significantly reduced periphery to whole-cell fluorescence ratio compared to the HA-tagged WT β-actin plasmid (Fig. 4 C, D; middle vs. upper panels). Consistently, while approximately 100% of the exogenous WT β-actin co-localized with endogenous β-actin, only approximately 10% of the mutant β-actin did so (Fig. 4E). This suggests impaired cellular localization and polymerization of the mutant β-actin. Furthermore, co-transfection of WT and mutant β-actin resulted in a decreased periphery to whole-cell fluorescence ratio for the WT β-actin (Fig. 4 F) and reduced co-localization with endogenous β-actin compared to WT-only transfected cells (Fig. 4G). These findings indicate a dominant-negative effect of the p.Gln360ProfsTer4 ACTB variant on WT β-actin function.
Impaired Immunological Synapse Formation and Reduced Intra-Synaptic IL-2 Concentration in T Cells with the ACTB Variantβ-actin, a key component of the actin cytoskeleton, plays a crucial role in T-cell activation by undergoing significant rearrangement. It contributes to the formation of the immune synapse and the molecular events during T-cell activation, ultimately impacting the cell’s ability to respond to antigenic stimuli and perform its immune functions [1]. Therefore, we hypothesized that T-cell activation and function in P1 are impaired. We first examined the formation of immunological synapses following activation. CD3+ T cells were isolated and activated for 16 h using Dynabeads™ Human T-Expander CD3/CD28. The T cells were then stained for CD3, actin, IL-2 and nucleus. Confocal microscopy analysis revealed defective immunological synapse formation upon activation in P1 as compared to HC, with reduced local intra-synaptic IL-2 concentrations in the patient (Fig. 5A, B). Noteworthy, the TCRα,β and CD28 expression in T cells from P1 and HCs was comparable (Supplementary Fig. 1). These findings suggest that T cells carrying the ACTB variant fail to form immunological synapses with antigen-presenting cells properly. Moreover, the reduced intra-synaptic IL-2 concentrations indicate impairment in signal transduction required for effective T-cell activation.
Fig. 5
Analysis of immunological synapse formation and IL-2 localization in T cells. (A) CD3+ T cells were isolated and activated for 16 h using Dynabeads™ Human T-Expander CD3/CD28. Cells were then stained with anti-CD3 (T-cell receptor complex), Phalloidin (F-actin), DAPI (nuclei), and anti-IL-2. Confocal microscopy was used to visualize the immunological synapse and assess IL-2 distribution. Comparisons were made between T cells from the patient (P1) and a healthy control (HC). (B) Quantification of percentages of synapse formation out of all T-cell-beads interactions. CD28/CD3-coated activating beads are marked with white arrows. Presented is a comparison between the patient and 3 healthy controls. The two-tailed, unpaired Mann–Whitney test was used for statistical analysis. Results are presented as mean ± SD (****P ≤ 0.0001)
The Dominant-Negative ACTB Variant Disrupts Activated T-Cell MorphologyAs immunological synapse formation and intra-synaptic IL-2 concentration were impaired in T-cell from P1, we next aimed to evaluate T-cell activation of P1 and to assess the effect of exogenous IL-2. Our group previously reported that activation and proliferation of CARMIL2-deficient T-cell could be rescued in-vitro by adding exogenous IL-2 [15]. Similarities between CARMIL2 deficiency and ACTB-related BRWS1 in P1, including T-cell cytoskeleton dysfunction and immune dysregulation, prompted us to attempt rescuing P1’s impaired T-cell effector functions by adding different doses of exogenous IL-2 to the activation medium.
Upon activation, T cells undergo a marked increase in size within the first 24 h [16], accompanied by extensive cytoskeletal remodeling [17], including actin polymerization [18] and elongation-flattening of the activated cells [19]. Additionally, during activation, T cells increase their nuclear size through chromatin de-condensation, which facilitates the upregulation of activation-related proteins [20]. To investigate these early events in T-cell response to activation, PBMCs from P1 and HC were activated for 24 h with or without the addition of exogenous IL-2 at different concentrations. After 24 h of activation, the cells were stained for actin (using phalloidin), CD3 and nucleus, and then visualized using confocal microscopy (Fig. 6A). The subsequent analysis focused on several morphological parameters, including surface area, maximal diameter, cellular perimeter, cellular elongation (the ratio of maximal to minimal diameter) and nuclear area of the activated T cells. Phalloidin signal intensity was also measured to quantify T-cell β-actin levels. In comparison to WT, P1 T cells demonstrated a significant reduction in surface area (Fig. 6B), nuclear area (Fig. 6 C), and β-actin levels (Fig. 6D). No significant alterations were observed between the two groups in maximal diameter (Fig. 6E), cellular perimeter (Fig. 6 F), and cellular elongation (Fig. 6G). These results indicate abnormal cell morphology and confirm β-actin deficiency in P1, consistent with the immunoblotting results (Fig. 3D). Furthermore, while exogenous IL-2 increased all these morphological parameters in a dose-dependent manner in HC’s activated T cells, it had no effect on the T cells carrying the p.Gln360ProfsTer4 ACTB variant (Fig. 6 H-L). However, there was no evidence that exogenous IL-2 increased the nuclear area in either group (Fig. 6M).
Fig. 6
Morphological analysis of activated T cells and the effect of exogenous IL-2. (A) PBMCs from the patient (P1) and healthy controls (HC) were activated for 24 h with or without the addition of exogenous IL-2. Cells were stained with phalloidin (F-actin), DAPI (nuclei), and anti-CD3 antibody to assess T-cell morphology. Confocal microscopy was used to visualize and quantify cellular and nuclear morphology. (B-G) Several morphological parameters were analyzed, including surface area, nuclear area, β-actin levels, maximal diameter, cellular perimeter, and cellular elongation. (H-M) The effect of exogenous IL-2 on these parameters was assessed in both HC and P1 T cells. Each dot represents an individual cell. The two-tailed, unpaired Mann–Whitney test was used for statistical analysis. Results are presented as mean ± SD (P values: ns = non-significant; *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001)
Impaired Activation and Proliferation of T Cells with the ACTB Variant, Partially Rescued In-Vitro by Exogenous IL-2Due to the lack of impact of exogenous IL-2 on morphological parameters of the patient’s T cells, we next aimed to examine its effect on effector functions. PBMC from the HC and the patient were activated in-vitro. After 48 h of activation, light microscopy revealed that T cells from P1 exhibited impaired formation of active T-cell rosettes, unlike those from HC (Supplementary Fig. 2). We then analyzed different T-cell effector functions using flow cytometry. Upregulation of CD25 (α subunit of IL-2 receptor) was reduced in P1’s effector CD4+ T cells compared to those of the HC (Fig. 7 A, B). Additionally, IFN-γ and IL-4 levels in the supernatants of 5-day-activated T cells were markedly decreased in P1 compared to the HC, indicating impaired Th1 and Th2 cytokine secretion, respectively (Fig. 7 C, D). Lastly, CD4+ T-cell proliferation capacity was also impaired, as demonstrated by CellTrace Violet (CTV) staining and flow-cytometry readout (Fig. 7E, F). Overall, these results demonstrated decreased activation and proliferation of CD4+ T cells in P1 with the dominant negative ACTB variant. Interestingly, a partial rescue was noted at higher IL-2 doses, as evidenced by CD25 upregulation (Fig. 7A-B), IFNγ and IL-4 secretion during activation (Fig. 7C-D), and CD4+ T-cell proliferation capacity (Fig. 7E-F). Measurements of secreted IL-4 and IFNγ following 5 days of activation revealed that the rescue of activation by addition of exogenous IL-2 was dose-dependent (Fig. 7C-D). Similar findings regarding decreased effector function and IL-2 partial rescue were observed for CD8+ T cells (Supplementary Fig. 3A-D). These findings are notably similar to our previous report demonstrating the rescue of activated CARMIL2-deficient T cells by adding exogenous IL-2 in-vitro [15]. However, despite the rescue in activation and proliferation, CD4+ and CD8+ T-cell post-activation counts were significantly reduced compared to the HC (supplementary Fig. 4A-C). Correspondingly, increased dead cell counts were noted in P1 (Supplementary Fig. 4D). In summary, these results demonstrate that T cells harboring the p.Gln360ProfsTer4 ACTB variant are characterized by impaired cellular morphology, activation and proliferation. While addition of exogenous IL-2 partially rescued activation and proliferation, it did not significantly affect the morphology of T cells harboring the p.Gln360ProfsTer4 ACTB variant.
Fig. 7
Analysis of T-cell activation, cytokine secretion, and proliferation in the presence of exogenous IL-2. (A-B) PBMCs from the patient and a healthy control were activated in vitro for 48 h, followed by flow cytometry analysis of CD25 (IL-2 receptor α) expression in CD4+ effector T cells. (C-D) Interferon (IFN)γ and IL-4 cytokine levels were measured in the supernatants of 5-day-activated T cells to assess Th1 and Th2 responses. Quantification of cytokines was conducted by ELISA (E-F) T-cell proliferation capacity was evaluated using CellTrace Violet (CTV) staining and flow cytometry. Presented are histograms of CTV staining. The effect of different doses of exogenous IL-2 on these parameters was also examined
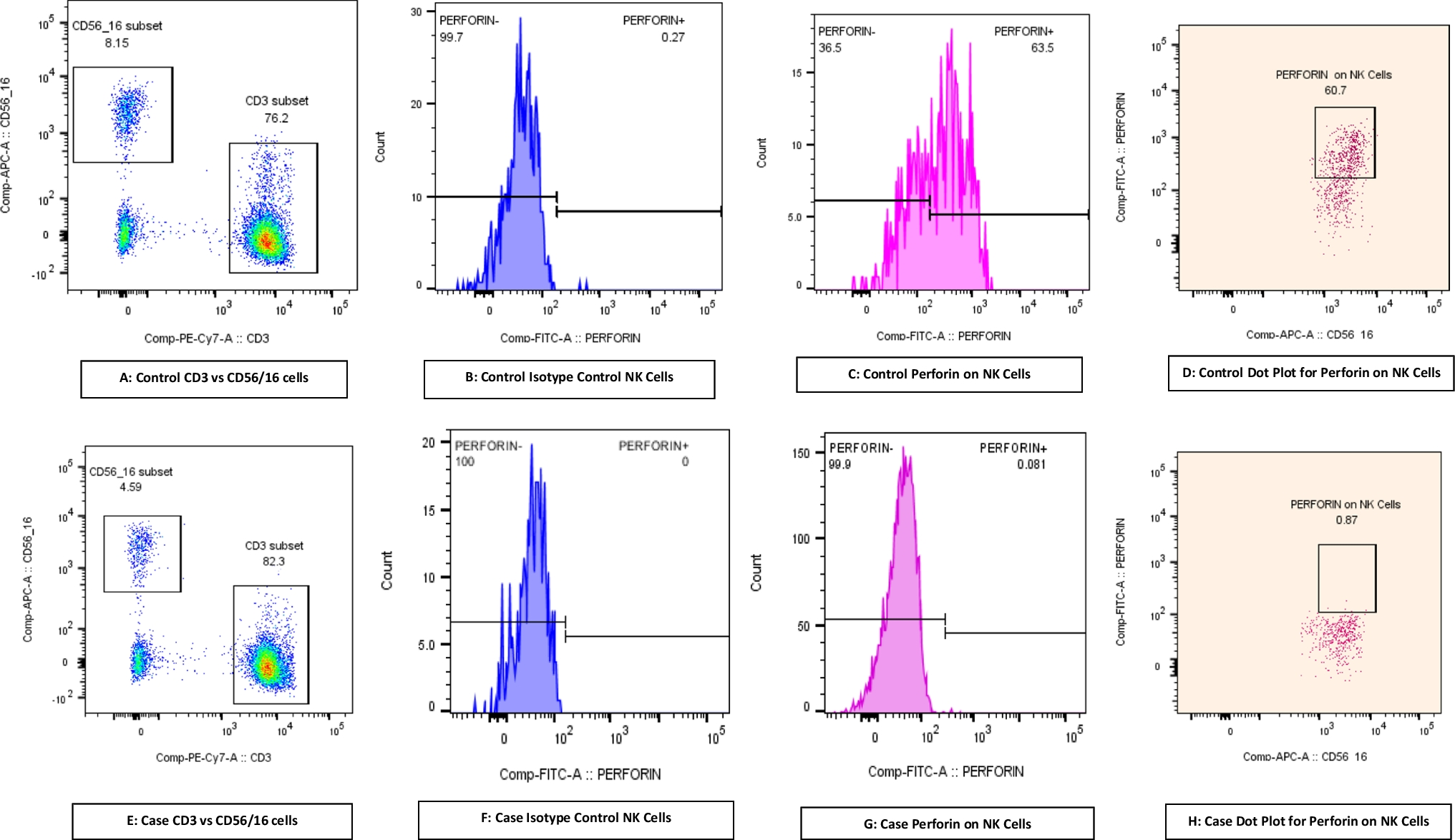
Immune Evaluation Reveals Specific Antibody Deficiency, Immune Dysregulation with Hyper-IgE Levels, and Significantly Reduced Memory T-Cell SubsetsUpon admission to our clinic, an immune evaluation was initiated (Table 1). A complete blood count indicated normal levels of absolute leukocytes, lymphocytes, and eosinophils; however, absolute monocyte counts were consistently elevated, with a value of 1.2 × 109/L (normal range: 0.28–0.5 × 109/L). Flow cytometry analysis showed an increased quantity of the naïve T-cell subset (TN) with a corresponding decrease in total memory (TM) and effector memory (TEM) CD4+ T-cell subsets in P1, as compared to HC (Fig. 8A-E). TEMRA CD4+ population was also reduced in P1 (Fig. 8G, H). Analysis of CD8+ T cells demonstrated similar results (Supplementary Fig. 5). B cell and NK cell counts were within normal limits. In terms of humoral immunity, hypergammaglobulinemia was observed, with increased levels of total IgG (1955 mg/dL; normal range: 639–1349 mg/dL), as well as IgG1 and IgG4 levels. Conversely, IgM levels were low at 25 mg/dL (normal range: 40–230 mg/dL). Extremely elevated IgE levels were detected, peaking at 16,700 U/mL (normal range: 0-100 U/mL). Furthermore, IgG titers against previous vaccines—including measles, mumps, rubella, varicella, hepatitis A and B, tetanus, pertussis, diphtheria, and Streptococcus pneumoniae—were all negative. This indicates a specific antibody deficiency despite elevated total IgG levels; therefore, the patient was advised to initiate monthly immune globulin replacement therapy (Table 1). Along with recurrent infections, P1 displayed atopy, including atopic dermatitis, asthma and allergic rhinitis. Consistent with immune dysregulation, P1 was found to have nearly absent circulating CD4+CD25+FOXP3+ (forkhead box protein 3) TREGS counts (0.96%; normal range: 4.2–9.9%). In addition, flow cytometry analysis of the TCR V-β repertoire revealed a normal, polyclonal distribution. However, a notable expansion of the Vβ3 clone was observed in the patient compared to HC, suggesting a potential autoreactive clonal expansion (Supplementary Table 1). Finally, since BRWS1 has been previously associated with neutrophil dysfunction [9], we aimed to assess reactive oxygen species (ROS) production in neutrophils using two methods: chemiluminescence and DHR assays. The results indicated that although P1’s neutrophils generated some ROS, both the total production and secretion in response to PMA activation were reduced in the patient compared to HC (Supplementary Fig. 6 A-B). Unfortunately, the patient declined the offered intravenous immunoglobulin and prophylactic antibiotic treatments, despite his history of recurrent infections and specific antibody deficiency. In summary, the immunological analysis of P1 indicated a phenotype of combined immune deficiency consistent with features of a primary immune regulatory disorder.
Table 1 Immune workup of the patient with dominant negative-ACTB variantFig. 8
Immune phenotyping analysis of T-cell subsets in the patient. PBMCs were purified from the patent’s peripheral blood, stained for surface markers and analyzed by flow cytometry for T-cell specific subsets. Data are presented as density plots and summarized in bar graphs. (A-F) Flow cytometric analysis of CD4+ T-cell subsets, including naïve (TN), total memory (TM), effector memory (TEM) and central memory (TCM) populations. (G-H) As explained above, for effector memory cells re-expressing CD45RA (TEMRA) CD4+ T-cell subset
Dupilumab Treatment for the Patient Successfully Alleviates Chronic InflammationTo address the patient’s immune dysregulation and symptoms consistent with type 2 inflammation, dupilumab, a monoclonal antibody targeting the IL-4/IL-13 receptor α subunit, was initiated at a dose of 300 mg every two weeks. Baseline serum cytokines were quantified in both the patient and HCs using ELISA, revealing elevated levels of Th1 cytokines (TNFα, IFN-γ), Th2 cytokines (IL-4, IL-13, IL-10), and pro-inflammatory cytokines (IL-6, IL-1β) (Fig. 9 A-E). Serum levels of IL-5 and IL-17 A were similar to those in HC, with IL-5 levels corresponding to the normal eosinophil counts observed in the patient (Fig. 9 C, D). During dupilumab treatment, the patient’s atopic dermatitis improved, as evidenced by reductions in Scoring Atopic Dermatitis (SCORAD) and Eczema Area and Severity Index (EASI) scores (Fig. 9 A). To objectively assess the impact of dupilumab, we measured various serum biomarkers at baseline (t = 0), and after 30 and 150 days of treatment. Dupilumab significantly reduced the Th1 cytokine IFN-γ, although TNFα levels remained unchanged (Fig. 9B). Th2-related cytokines, including IL-13, IL-5 and IL-10, as well as IgE, also decreased over time, as anticipated (Fig. 9 C). Interestingly, serum IL-4 levels increased, which may reflect a compensatory response or serum accumulation due to IL-4 receptor blockade by dupilumab (Fig. 9 C). IL-17 A levels remained stable, with a slight increase observed during treatment (Fig. 9D). Evidence of remission from chronic general inflammation was observed through reduced serum levels of the pro-inflammatory cytokines IL-1β and IL-6 (Fig. 9E), as well as C-reactive protein (Fig. 9 F). Additionally, blood count-derived inflammatory markers have been shown to be useful for monitoring during dupilumab treatment [21]. Specifically, the platelet-to-lymphocyte ratio (PLR), monocyte-to-lymphocyte ratio (MLR), neutrophil-to-lymphocyte ratio (NLR), systemic inflammation response index (SIRI), and systemic immune-inflammation index (SII) all decreased during treatment with dupilumab in our patient, again indicating resolution of overall systemic inflammation. Notably, the percentage of circulating TREGS increased from a baseline level of 0.96–2.18% (Fig. 9H, I), which may indicate a partial resolution of immune dysregulation induced by dupilumab. In summary, these findings suggest that dupilumab treatment in the patient with the ACTB variant effectively managed atopic dermatitis, while also reducing overall inflammation and potentially shifting immune responses by suppressing both Th1 and Th2 pathways, while preserving Th17-mediated immunity.
Fig. 9
Clinical and Immunological responses to dupilumab treatment. (A) Eczema area and severity index (EASI) and Scoring atopic dermatitis (SCOARD) clinical scores of the patient’s atopic dermatitis during dupilumab treatment. (B-F) Longitudinal analysis of serum Th1, Th2, Th17 and pro-inflammatory cytokines, IgE levels, absolute eosinophil count and C-reactive protein at baseline (t = 0), and after 30 and 150 days of dupilumab treatment. (G) Blood count-derived inflammatory markers during dupilumab treatment, including the platelet-to-lymphocyte ratio (PLR), monocyte-to-lymphocyte ratio (MLR), neutrophil-to-lymphocyte ratio (NLR), systemic inflammation response index (SIRI), and systemic immune-inflammation index (SII). (H-I) Flow cytometric analysis of circulating regulatory T cells (TREGS) at baseline and during treatment with dupilumab
Comments (0)