
Remember me



The patient, a male, was admitted to Fudan University Children’s Hospital at the age of 10. Beginning at the age of 5, he developed a recurrent cough with yellow purulent sputum and intermittent fever, requiring anti-infective therapy 1–2 times a month. Multiple CT scans of the lungs revealed pulmonary infections and bronchiectasis. The patient underwent multiple bronchoscopy procedures, and the pathogens in the alveolar lavage fluid included Mycoplasma, Haemophilus influenzae, Pseudomonas aeruginosa, and Staphylococcus aureus. An increase in galactomannan (GM) in the alveolar lavage fluid was recorded (4.917), indicating the presence of a fungal infection. Despite treatment with a variety of antibacterial and antifungal drugs, symptoms of cough and expectoration persisted, and the patient eventually exhibited abnormal ventilatory function. In addition to recurring pneumonia, he had a history of thrush, eczema, and viral warts, and an MRI of the head suggested otitis media and sinusitis. The levels of cancer antigen 199 (CA199) and CA125 were significantly elevated, but no neoplastic lesions were found on abdominal CT. At 12 years old, he developed swollen cervical and supraclavicular lymph nodes with tenderness, and a PET-CT suggested reactive hyperplasia. Cytomegalovirus (CMV) was detected in the blood, urine, and sputum, and antiviral therapy with ganciclovir was subsequently initiated. The patient is currently 13 years old, with a height of 152 cm (10th–25th percentile) and a weight of 33.6 kg (below the 10th percentile). He continues to experience recurrent cough and sputum production, requiring intermittent anti-infective therapy. IVIG is not currently being administered. Human leukocyte antigen (HLA) typing has been completed, and hematopoietic stem cell transplantation (HSCT) is planned for the future.
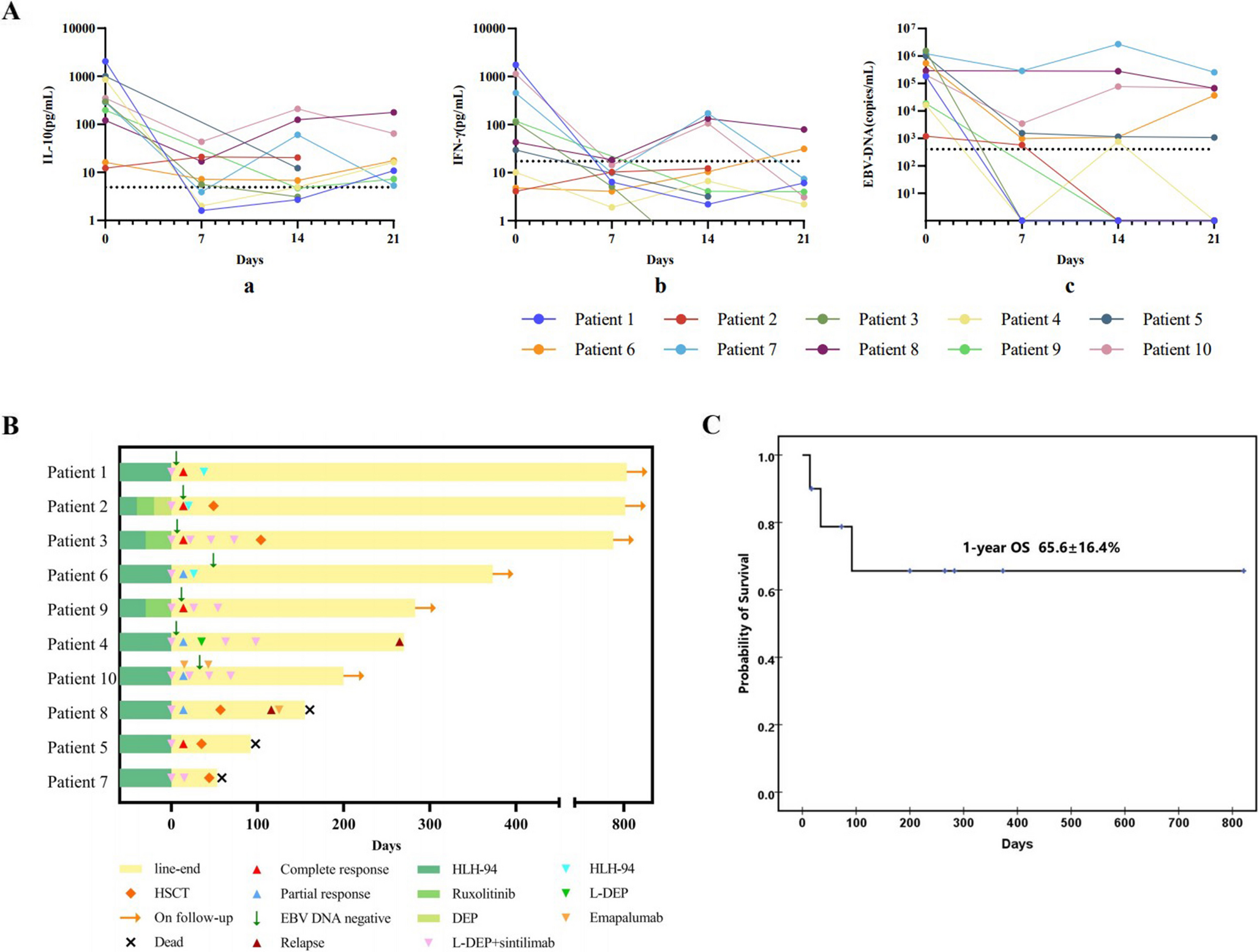
ImmunophenotypingRoutine blood examinations and flow cytometric analysis of peripheral blood for immune cell subsets revealed that the patient had normal counts of T, B, and NK cells (Table 1). The patient had an increased proportion of naïve B cells, but significantly reduced proportions of memory B cells and switched memory B cells. He exhibited a decreased proportion of CD4+ T cells and an increased proportion of CD8+ T cells, resulting in an inverted CD4/CD8 ratio, with a decreased proportion of naïve T cells and a markedly increased proportion of memory T cells (Table 2). Serum levels of immunoglobulins (Ig) G, A, M and E were increased, with IgE levels rising to 5503 KU/L. Allergen testing indicated positive reactions to egg whites (15.9 KU/L) and fungi (2.32 KU/L). Despite receiving hepatitis B vaccination at birth, the patient tested negative for hepatitis B surface antibody at 11 years of age. The TCR and BCR repertoires of the patient did not differ significantly from those of HC (Fig. S1).
Table 1 Routine blood tests for the patientTable 2 Flow cytometric analysis of immune cell subsets in the peripheral blood of the patientGenetic AnalysisWES was conducted on genomic DNA extracted from the peripheral blood of the patient and his family members. A missense variant, c.420 A > T (p.R140S), was identified in the patient, whereas his mother and sister were heterozygous for this variant (Fig. 1A). No other known genetic alterations that could account for the patient’s clinical phenotype were identified. Unexpectedly, Sanger sequencing revealed the presence of both IL2RG c.420 A > T and c.420 A > G variants in the patient’s peripheral blood gDNA (Fig. 1B); the latter, a silent mutation, was not detected in the WES data. TA cloning demonstrated that 39 out of 47 sequences (82.98%) from the patient’s peripheral blood gDNA were c.420 A > T, while 8 out of 47 sequences (17.02%) were c.420 A > G (Fig. 1B). However, only c.420 A > T was identified in the gDNA from the patient’s oral mucosa cells (Fig. 1C). Given the localization of the IL2RG gene on the X chromosome, it is probable that the patient inherited the 420T allele from his mother, similar to his sister. The 420G allele likely represents a revertant mutation that occurred in immune cells. Although WES confirmed a normal 46XY karyotype in the patient, maternal engraftment, particularly microchmerism, cannot be completely excluded due to the limited sensitivity of karyotyping. The proportions of the 420T allele and the 420G reversion varied among different lymphocyte subpopulations (Fig. 1D). Specifically, 0 of 20 sequences (0%) in B cells were 420G, while 1 of 11 (9.09%) in CD4+ T cells and 3 of 23 (13.04%) in CD8+ T cells were 420G. In NK cells, 6 of 14 (42.86%) sequences were 420G. The differential proportions of the 420G revertant among various immune cells likely reflect selective pressure during immune cell development and function. The p.R140S variant is localized in the extracellular domain of γc (Fig. S2), and is predicted to be deleterious by in silico algorithms (Fig. 1E). Based on these findings, we proceeded with a more detailed investigation of this variant.
Fig. 1
Verification of the variant and identification of a revertant mutation. (A) Family pedigree of the patient. (B) Sequence profiles of PCR products from HC genomic DNA (gDNA), the patient’s gDNA, and monoclonal colonies following ligation of the patient’s PCR products into the T vector. (C) Sanger sequencing of gDNA amplification products from the patient’s oral mucosa cells. (D) Analysis of mutation and reversion rates in different lymphocyte subpopulations. (E) In silico predictions of variant’s deleteriousness
The c.420 A > T (p.R140S) Variant Impairs γc ExpressionTo evaluate the impact of the R140S amino acid change on γc expression, we used flow cytometry to analyze both cell surface and total (cell surface + intracellular) γc expression in various immune cell subsets before and after α-CD3 + α-CD28 stimulation (Fig. 2). In CD19+ B cells, both cell surface and total γc expression levels were lower in the patient compared to HCs, both before and after stimulation (Fig. 2A and B, left column). In the patient’s CD4+ and CD8+ T cells, a bimodal distribution of γc expression was observed (Fig. 2A and B, 2nd and 3rd columns), consistent with the presence of revertant variant in these cells. The higher γc expression peak was comparable to that of HCs, while the lower peak was notably reduced. Additionally, γc expression levels in NK cells were only slightly lower than those in HCs (Fig. 2A and B, right column), which corresponds with the high proportion of the revertant variant in these cells. Overall, γc protein expression was severely diminished in B cells, substantially reduced in T cells, and moderately decreased in NK cells, correlating with the proportions of the revertant variant in these immune cell subsets.
Fig. 2
γc expression in the patient’s immune cells. Surface and total (surface + intracellular) γc staining in gated CD19+ B cells, CD4+ T cells, CD8+ T cells, and CD56+ NK cells, before (A) and after (B) α-CD3 + α-CD28 stimulation for 3 days. MFI values for γc under each condition are shown
Reduced Phosphorylation of STAT3 and STAT5 in the Patient’s Immune CellsTo determine whether the variant affects IL-21/IL-7/IL-2 receptor signaling, we analyzed the phosphorylation of STAT3 and STAT5 following 10-minute stimulations with IL-21, IL-7 and IL-2, respectively. Before stimulation, the patient’s B cells showed p-STAT3 levels comparable to those of three HCs. However, these cells demonstrated reduced upregulation of p-STAT3 after IL-21 stimulation compared to HCs (Fig. 3A, top panels). Similarly, the patient’s CD19−CD4−CD8− cells, primarily NK cells and monocytes, also showed p-STAT3 levels comparable to HCs before stimulation, but displayed reduced upregulation after IL-21 stimulation (Fig. 3A, bottom panels). In contrast, both CD4+ T and CD8+ T cells from the patient had p-STAT3 levels similar to HCs before stimulation and exhibited a similar increase in p-STAT3 levels following IL-21 stimulations (Fig. 3A, 2nd and 3rd rows).
Fig. 3
Phosphorylation levels of STAT3 and STAT5 following IL-21 and IL-7 stimulation. (A) Levels of phosphorylated STAT3 (p-STAT3) and (B) phosphorylated STAT5 (p-STAT5) in gated CD19+ B cells, CD4+ T cells, CD8+ T cells, and CD19−CD4−CD8− cells, measured before (red) and after (blue) a 10 min stimulation with 10 ng/mL IL-21 (A) and 10 ng/mL IL-7 (B)
For p-STAT5, B cells from both the patient and HCs showed comparable levels prior to stimulation, with only marginal upregulation following IL-7 exposure (Fig. 3B, top panels). Similarly, the CD19−CD4−CD8− cells from the patient and HCs had comparable p-STAT5 levels before stimulation. After IL-7 stimulation, a fraction of CD19−CD4−CD8− cells in HCs demonstrated increased pSTAT5 levels, although the proportion of responsive cells varied among individuals, with HC3 showing the lowest proportion. In contrast, the patient’s CD19−CD4−CD8− cells displayed minimal upregulation of p-STAT5 levels and lacked a prominent responsive fraction (Fig. 3B, bottom panels). Unlike B cells, CD4+ T and CD8+ T cells from the patient had p-STAT5 levels comparable to HCs before stimulation but demonstrated significantly impaired upregulation of p-STAT5 levels after IL-7 stimulation (Fig. 3B, 2nd and 3rd rows). In addition to impaired responses to IL-21 and IL-7, the patient’s CD4+ T cells, CD8+ T cells, and CD19−CD4−CD8− cells also exhibited significantly reduced pSTAT5 upregulation following IL-2 stimulation (Fig. S3A). Collectively, these results indicate that the R140S variant impairs γc-dependent signaling, resulting in diminished IL-21 responses in B cells and NK cells, and reduced IL-7 and IL-2 signaling in CD4+ T cells, CD8+ T cells, and NK cells.
Correlation of p-STAT3 and p-STAT5 Levels Induced by IL-21 and IL-7 With Cell Surface γc ExpressionThe γcR140S variant exhibited reduced protein expression and diminished levels of p-STAT3 and p-STAT5. To further investigate the relationship between γc expression levels and signaling effects, we analyzed mean fluorescence intensity (MFI) of p-STAT3 and p-STAT5 relative to γc expression. As illustrated in Fig. 4A, p-STAT3 levels correlated with cell surface γc expression following IL-21 stimulation. A similar pattern was observed for p-STAT5 following IL-7 stimulation, as depicted in Fig. 4B. This correlation was particularly pronounced in the patient’s CD4+ and CD8+ cells, where a fraction of cells that restored γc expression through the revertant mutation displayed relatively higher p-STAT5 levels (Fig. 4B). These findings suggest that the R140S variant primarily impairs γc function by reducing its expression on the cell surface.
Fig. 4
Correlation of STAT phosphorylation with γc surface levels. (A) p-STAT3 and (B) p-STAT5 levels in gated CD19+ B cells, CD4+ T cells, CD8+ T cells, and CD19−CD4−CD8− cells following a 10-minute stimulation with 10 ng/mL IL-21 (A) and 10 ng/mL IL-7 (B). The X-axis displays γc levels, and the Y axis shows p-STAT levels
γc Variant Impairs T and B Lymphocyte ProliferationTo assess the impact of the γcR140S variant on lymphocyte proliferation, we labeled fresh PBMCs with CFSE and analyzed CFSE dilution in gated CD19+ B cells, CD4+ T cells and CD8+ T cells following various stimuli using flow cytometry. A portion of the CD19+ B cells from HCs underwent cell division in response to CpG, CpG + IL-2, α-CD40, and α-CD40 + IL-4 + IL-21 (Fig. 5A). In contrast, the patient’s CD19+ B cells showed minimal division under the same conditions. Notably, HC2 showed a similarly poor response to α-IgM stimulation as the patient. In response to IL-7, both CD4+ and CD8+ T cells from HCs underwent multiple rounds of cell division (Fig. 5B); however, only a small fraction of the patient’s CD4+ and CD8+ T cells exhibited limited proliferation. Nonetheless, the patient’s CD4+ T and CD8+ T cells displayed substantial cell division comparable to HCs in response to α-CD3 + α-CD28 stimulation (Fig. 5).
Fig. 5
Analysis of B and T cell proliferation. CFSE-labeled PBMCs from HCs and the patient were cultured with various stimuli for 6 days and analyzed for CFSE dilution in gated CD19+ (A), CD4+ T (B), and CD8+ T (C) cells. The proportions of divided and non-divided cells are shown in the left and right of each panel, respectively
We further used frozen PBMCs of the patient and two HCs and assessed T cell proliferation in response to IL-2 and IL-15, using IL-7 responses as controls. Although cytokine responses in frozen PBMCs were generally weaker than those observed in fresh samples, the patient’s CD4+ and CD8+ T cells both showed a lower proportion of divided cells in response to IL-7 compared to HCs and moreover lacked the subset of cells that had undergone multiple rounds of division, which was present in HCs (Fig. S3B-C). Similarly, the patient’s CD4+ and CD8+ T cells exhibited reduced proliferation in response to IL-2 and IL-15 (Fig. S3B-C). These results indicate that the γcR140S variant impairs B cell proliferation in response to multiple stimuli (except α-IgM) and reduces T cell proliferation in response to IL-7, IL-2, and IL-15, while T cell proliferation induced by α-CD3 + α-CD28 stimulation remained comparable to that of HCs.
Structure of the IL-7-IL-7RA-γc Complex is not Affected by R140S Amino Acid ChangeTo explore the mechanisms behind the reduced protein expression of the γcR140S variant, we first employed the AlphaFold3 algorithm to assess the impact of the γcR140S variant on the structural configuration of the IL-7-IL-7RA-γc complex. As illustrated in Fig. S4A, R140 of γc is located away from the interaction interface with IL-7 or IL-7RA and does not affect hydrogen bond formation between the proteins. The irregularly curled region depicted in Fig. S4B represents a zone where AlphaFold3 exhibits suboptimal predictive performance [25]. Despite this, WT γc and γcR140S demonstrated a high degree of structural similarity. R140 is located in a curly region between β-Sheets, which does not affect the structure of γc itself (Fig. S4C). Therefore, the R140S amino acid substitution is unlikely to affect the interaction between γc and IL-7RA or the binding of IL-7 to the IL-7 receptor. However, the mutation of the basic amino acid R to the neutral amino acid S may reduce the basicity of γc, potentially resulting in abnormal localization of γcR140S.
Aberrant Localization of γcR140STo investigate the causes of decreased γc expression in the patient’s lymphocytes, we established Jurkat cells expressing γc-EGFP fusion proteins. We observed that WT γc localized in punctate clusters, whereas γcR140S displayed a disperse distribution (Fig. 6). To further substantiate the localization of WT γc and γcR140S within living cells, Jurkat cells expressing the γc-EGFP fusion protein were incubated with a series of probes that label acidic compartments, including lysosomes and late endosome, the ER, and the Golgi apparatus [23, 26]. WT γc primarily co-localized with acidic compartments (Fig. 6A) and also showed some co-localization with the ER (Fig. 6B), whereas γcR140S predominantly co-localized with the ER (Fig. 6B). Neither WT γc nor γcR140S exhibited significant co-localization with the Golgi (Fig. 6C).
Fig. 6
Aberrant subcellular localization of γcR140S. Confocal microscope images of Jurkat cells expressing WT γc-EGFP or γcR140S-EGFP fusion proteins after incubation with Lyso-tracker (A, magenta), ER-tracker (B, red), Golgi-tracker (C, red). Bright-field images also captured
R140S Variant does not Affect the Half-Life or Degradation Pathways of γcTo investigate whether aberrant protein localization affects the stability and degradation of γcR140S, we used Jurkat cells recently transduced with retroviruses expressing either WT γc-FLAG or γcR140S-FLAG fusion proteins and treated them with various inhibitors. The half-life of the proteins, assessed using the protein synthesis inhibitor cycloheximide (CHX), revealed no differences between WT γc and γcR140S (Fig. S5A). To analyze the degradation pathways of γc, we treated cells with different inhibitors. As depicted in Fig. S5B, the degradation of both WT γc and γcR140S was inhibited by NH₄Cl, a lysosomal inhibitor, but not by 3-MA (3-methyladenine), an autophagy inhibitor. We then cultured Jurkat cells recently transduced with retroviruses expressing these fusion proteins for four months and repeated the above experiments. This time, we also included MG132, a proteasome inhibitor. Again, we found no differences in the half-life (Fig. S5C) or degradation pathways (Fig. S5D) between WT γc and γcR140S. These findings suggest that both WT γc and γcR140S are primarily degraded via the lysosomal pathway, consistent with previous studies [27], and not via autophagy or proteasome pathways. Intriguingly, after four months of culture, the protein levels of γcR140S were significantly lower than those of WT γc (Fig. S5C, 0 h and Fig. S5D, Medium). These observations prompted us to examine whether γcR140S might induce or enhance an unfolded protein response (UPR) resulting in apoptosis in cells expressing high levels of γcR140S. We employed thapsigargin (TG) to induce the UPR in Jurkat cells expressing WT γc-FLAG or γcR140S-FLAG fusion proteins and analyzed the temporal changes in the protein levels of γc-FLAG, XBP1s, and BIP (Fig. S6A). Upon UPR induction, protein levels of both WT γc and γcR140S decreased similarly (Fig. S6B), accompanied by comparable increases in the expression of UPR signature proteins, such as XBP1s (Fig. S6C) and BIP (Fig. S6D). Therefore, γcR140S does not induce a stronger UPR compared to WT γc in Jurkat T cells.
Jurkat Cells Expressing High Levels of γcR140S Undergo Counterselection During Cell CultureIn Jurkat cells recently transduced with retroviruses expressing WT γc-EGFP or γcR140S-EGFP fusion proteins, the MFI of EGFP was initially similar but decreased at different rates over time during cell culture. After one month, the MFI for WT γc-EGFP decreased to 75%, whereas that for γcR140S-EGFP dropped to 54% (Fig. 7A). These findings suggest that Jurkat cells expressing high levels of γcR140S-EGFP are at a competitive disadvantage in cell culture. Moreover, in Jurkat cells expressing WT γc-FLAG or γcR140S-FLAG fusion proteins, the ratio of γcR140S-FLAG relative to WT γc-FLAG protein levels, as measured by immunoblot, progressively declined during culture, from 1.28-fold to 0.45-fold (Fig. 7B), suggesting a counterselection against cells with high levels of γcR140S expression.
Fig. 7
Counterselection of Jurkat cells expressing high levels of γcR140S in cell culture. (A) FACS analysis showing the EGFP MFI in Jurkat cells expressing WT γc-EGFP vs. γcR140S-EGFP from early (Day 0) to later (Day 31) stages in culture. (B) Comparison of γcR140S to WT γc expression over time in Jurkat cells expressing WT γc-FLAG vs. γcR140S-FLAG, analyzed by immunoblot with an α-FLAG antibody. The ratios of γcR140S-FLAG to WT γc-FLAG protein at each time point (Day 0, Day 10, Day 34, Day 119) are presented
Comments (0)