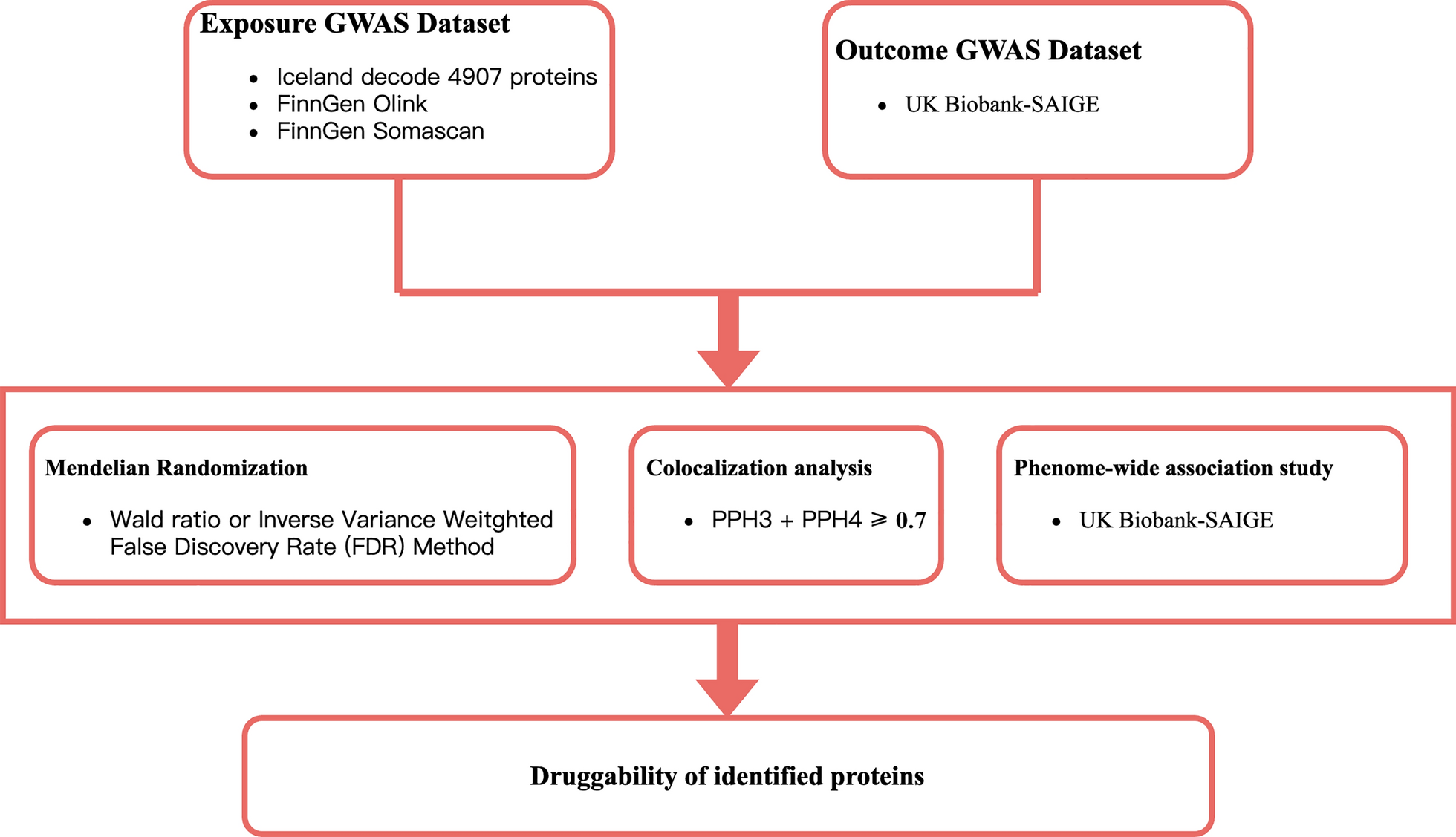
The treatment of CRC, particularly in its advanced stages, faces significant limitations despite the use of multimodal therapies, including surgery, chemotherapy, and targeted therapy. In our study, we explored the potential roles of GREM1, DKKL1, CHRDL2, and TMEM132 A as key proteins implicated in CRC pathogenesis, using Phe-WAS Mendelian randomization (PWMR). Our results suggest that these proteins, through their involvement in critical pathways such as Wnt signaling and BMP antagonism, may serve as novel therapeutic targets for CRC. This section will explore the significance of these findings in the context of recent research and discuss the therapeutic potential and challenges associated with targeting these proteins.
GREM1 plays a dual role in cancer biology by antagonizing BMP signaling, which is known to suppress tumor growth and promote cell differentiation in the gastrointestinal tract [22, 23]. In CRC, GREM1 overexpression has been shown to disrupt BMP-mediated tumor suppression, thereby allowing Wnt signaling to dominate and drive tumor progression. Studies have demonstrated that BMP antagonists like GREM1 are associated with increased cellular proliferation, resistance to differentiation, and enhanced invasive potential in CRC [24]. Our study reinforces these findings, showing that elevated GREM1 expression correlates with more aggressive CRC phenotypes. When compared with other studies, such as Gao, Z et al. [23], who explored BMP antagonists in CRC progression, the results align with the concept that restoring BMP signaling through GREM1 inhibition could reverse tumor growth and enhance the differentiation of cancer cells. Additionally, this protein’s involvement in Wnt signaling suggests a strategic point for therapeutic intervention, as targeting GREM1 may simultaneously modulate multiple tumor-promoting pathways. However, as highlighted in recent literature, targeting GREM1 could result in unintended effects on normal tissue homeostasis, particularly in organs where BMP signaling plays a critical role, such as the intestine [25].
DKKL1, a member of the Dickkopf glycoprotein family, functions primarily as an antagonist of the Wnt/β-catenin signaling pathway, which plays a key role in cell growth and differentiation. Dysregulation of this pathway is a hallmark of CRC, where DKKL1 not only promotes tumor proliferation and invasion but also modulates the tumor microenvironment by influencing immune cell activity [26, 27]. Our Mendelian randomization analysis identified genetically predicted DKKL1 levels as positively associated with CRC risk, consistent with previous findings suggesting a potential role of DKKL1 in oncogenic Wnt signaling. Given its dual role in Wnt regulation and immune modulation, DKKL1 has emerged as a promising therapeutic target, particularly for combination therapies involving immunotherapy [28]. Studies have shown that anti-DKKL1 therapies can synergize with immune checkpoint inhibitors to enhance CD8+ T-cell activity while reducing myeloid-derived suppressor cells (MDSCs), thereby overcoming tumor immune evasion [26, 29]. Our findings support the potential of DKKL1 as a direct therapeutic target, with colocalization analysis revealing shared genetic variation between DKKL1 and CRC, and phenotype-wide association studies showing no significant side effects linked to DKKL1 inhibition. Future research should focus on the long-term effects of DKKL1 inhibition and its integration with immunotherapies, particularly for patients with microsatellite-stable CRC, who typically exhibit poor responses to current treatments.
CHRDL2 (Chordin-like protein 2), a BMP antagonist, plays a key role in modulating BMP signaling, which is essential for maintaining cellular differentiation and tissue homeostasis. Dysregulation of BMP signaling, particularly through CHRDL2-mediated inhibition of BMP-4, can disrupt its tumor-suppressive effects, thereby promoting CRC progression [30]. Our MR analysis identified CHRDL2 as genetically associated with increased CRC risk (P_fdr < 0.05), with colocalization analysis (PPH3 + PPH4 > 0.7) further supporting its role as a direct therapeutic target. Given its ability to inhibit BMP signaling, targeting CHRDL2 could restore normal cellular differentiation, reducing tumor proliferation and enhancing therapeutic outcomes [30]. Prior research has linked BMP pathway disruption to increased tumor aggressiveness, although clinical studies specifically addressing CHRDL2 inhibition are limited to preclinical models. Our study strengthens the genetic basis for CHRDL2's involvement in CRC, showing no significant side effects (P_fdr > 0.05) and positioning it as a viable target for therapeutic intervention. Future research should focus on developing CHRDL2-specific inhibitors and evaluating their efficacy in vivo models, potentially combining these with immunotherapy or Wnt pathway inhibitors to maximize therapeutic benefit and overcome resistance mechanisms.
In contrast to the positive associations of genetically predicted GREM1, DKKL1, and CHRDL2 levels with CRC risk, our study suggests that TMEM132 A may have a potential protective role. The TMEM (Transmembrane Protein) family comprises various proteins with largely unexplored functions, particularly in cancer biology. Depending on the specific protein, TMEM members can either promote or inhibit tumorigenesis [31]. While TMEM132 A has been primarily studied in neurobiology, its role in CRC has not been extensively examined until now [32, 33]. Our study provides exploratory evidence that TMEM132 A acts as a protective factor in colorectal cancer (CRC), marking a novel association not previously identified in the literature. The findings suggest that higher TMEM132 A expression correlates with reduced tumor growth and invasiveness, indicating a potential tumor-suppressive role through its regulation of cell adhesion and migration. This aligns with prior studies showing high TMEM132 A expression correlates with better prognostic outcomes in other cancers, suggesting that its protective effects may extend across malignancies [32]. While promising, these results are preliminary and warrant further investigation to validate TMEM132 A’s role and elucidate its mechanisms, particularly its interactions with critical pathways such as Wnt signaling. Additional studies are necessary to confirm the therapeutic potential of targeting TMEM132 A in CRC.
The integration of PWMR with proteomic analysis provides a powerful approach for uncovering causal relationships between protein expression and CRC risk. By leveraging genetic data, PWMR enables the identification of protein biomarkers that may serve as therapeutic targets, while proteomics offers detailed insights into their functional roles within the tumor microenvironment. Our study demonstrates that combining these methodologies can reveal critical proteins, such as GREM1, DKKL1, CHRDL2, and TMEM132 A, that are causally linked to CRC progression. Further Phe-WAS did not reveal any significant adverse phenotypic associations for these targets (all P_FDR > 0.05). These plasma proteins, particularly GREM1 and CHRDL2, may offer novel entry points for therapeutic development. While their integration into current CRC treatment paradigms remains to be clarified, our findings provide foundational data for future mechanistic and translational efforts.
Recent advances in multi-omics integration have emphasized the value of combining proteomic, genomic, and transcriptomic data to gain a systems-level understanding of CRC biology. Integrative efforts, such as coupling CRISPR-Cas9 functional screens with quantitative proteomics, have revealed protein-level drivers of therapeutic resistance in CRC, providing new leads for drug development and mechanistic insights into tumor progression [34]. These technologies, together with Phe-WAS, allow for the interrogation of broader phenotypic effects. In our analysis, while some associations with tumor-related phenotypes were observed, no major safety concerns emerged, supporting the translational viability of the candidate targets.
In this study, we identify GREM1, DKKL1, CHRDL2, and TMEM132 A as plasma proteins causally associated with CRC risk, implicating them in key axes of Wnt/BMP signaling, immune modulation, and redox homeostasis. Notably, GREM1 and CHRDL2, as BMP antagonists, may potentiate Wnt-driven epithelial proliferation by suppressing BMP-mediated differentiation cues [35, 36]. GREM1, in particular, has also been linked to stromal remodeling and immunosuppression in glioblastoma and pancreatic cancer, suggesting conserved oncogenic functions across tumor types [37]. DKKL1, a non-canonical Wnt modulator, intersects with immune-metabolic regulation and has been associated with immune evasion mechanisms in myeloma and prostate cancer [38, 39]. Conversely, TMEM132 A appears negatively correlated with CRC risk, possibly through modulation of redox signaling and innate immunity, consistent with tumor-suppressive roles described in lung and renal cancers [32, 40]. Together, these findings highlight proteomic links between epithelial signaling, immune evasion, and metabolic stress, situating these proteins within a broader oncogenic context. Although our Phe-WAS revealed no major off-target liabilities, intertumoral heterogeneity in lipid metabolism [41] and immune architecture [42] underscores the need for future studies integrating proteogenomic, immune, and metabolic stratification to refine therapeutic utility. These findings point to the necessity for integrated multi-omics approaches, not only to validate these candidates at the functional level, but also to address potential redundancy within signaling pathways and refine therapeutic targeting strategies accordingly.
While our study provides valuable insights into the roles of GREM1, DKKL1, CHRDL2, and TMEM132 A in CRC, several limitations must be acknowledged. First, our findings are based on genetic associations and in silico analyses, which cannot definitively establish causality. The protein-trait relationships require further validation through functional experiments. Potential constraints include population stratification, database limitations, and the complex nature of protein interactions, which may influence the generalizability of our results. While MR-Egger and weighted median methods were employed to mitigate horizontal pleiotropy, residual bias may persist due to unmeasured pleiotropic pathways. Future studies should incorporate multivariable MR to adjust for shared genetic confounding. Another limitation of our study is the limited ancestral diversity of the cohort, which is primarily composed of individuals of European descent. This homogeneity may affect the trans-ethnic portability of our findings, especially given that plasma protein levels, genetic regulation, and tumor-immune interactions can vary significantly across populations. Future studies incorporating diverse ancestry groups will be critical to validate these associations and uncover population-specific molecular drivers. Additionally, the reliance on summary-level genetic data and proteomics databases means that individual variability and potential technical biases cannot be fully excluded. These limitations underscore the need for future prospective studies and experimental validation to confirm our computational findings. Further studies should employ comprehensive proteomic screens and functional assays to validate these findings and explore the broader proteomic landscape of CRC and also investigate the potential for combination therapies that target these proteins in conjunction with established treatments like chemotherapy, immunotherapy, and radiation. Such approaches may enhance treatment efficacy and overcome resistance mechanisms, ultimately improving outcomes for patients with advanced CRC.





Comments (0)