
Remember me
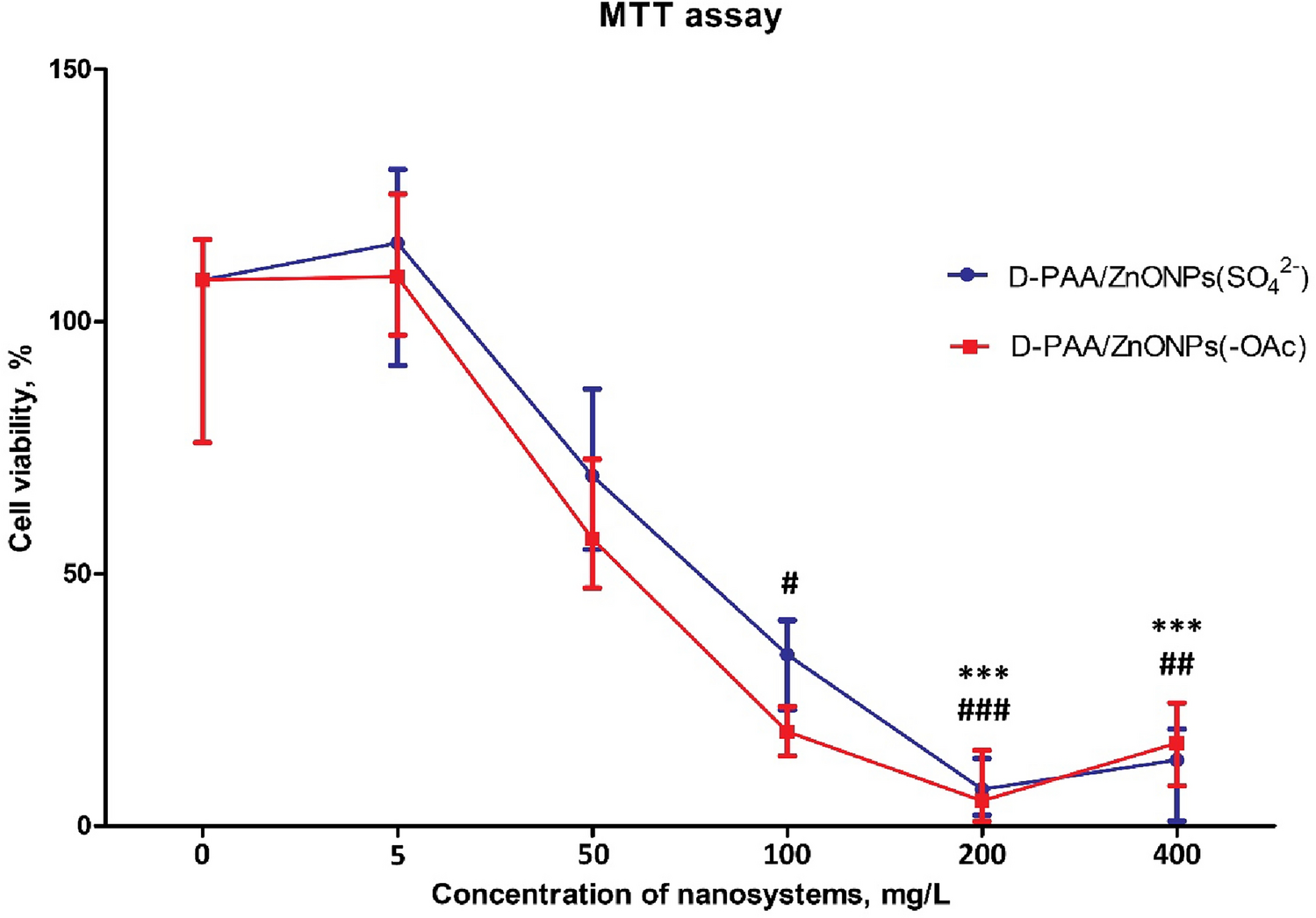


As a control, we first measured the isotopic composition of uninfected E. coli cells grown in minimal medium, by EA-IRMS (Fig. 1). The sole carbon source was d-glucose containing various proportions of 13C6-d-glucose. Overall, the measured 13C content of cells was slightly inferior to the theoretical 13C content of the substrate, with the relative difference varying between − 13.55 and 1.40%. Such difference cannot be explained by the addition of the inoculum, which contained LB medium, because a 1000× dilution was applied for inoculation. It could rather result from the 13C content of 13C6-d-glucose being slightly inferior to 100% (≥ 99% according to the supplier), combined with biases introduced during medium preparation and IRMS measurement. Despite these minor differences, a very strong correlation was obtained between the 13C content of substrate and cells, as expected (Fig. 1, R2 = 0.999). The replicates (N = 2) showed high reproducibility.
Fig. 1
Isotopic composition of E. coli cells, as determined by EA-IRMS, in function of the percentage of 13C6-d-glucose employed in the d-glucose substrate. There are 4 replicates by condition. The different shapes, circles, and triangles represent two distinct experimental series. The dotted line and the equation shown in the plot are related to the linear regression
The buoyant Density of T4 DNA Correlates with the Isotopic Composition of the Substrate Used for Host GrowthSubsequently, E. coli cells grown on minimal medium with various proportions of 13C6-d-glucose were infected by T4 bacteriophage. T4 DNA was extracted and separated on a CsCl gradient. A good reproducibility was observed among replicates, and small differences are visible across the two different experimental series. Overall, a strong correlation was obtained between the observed T4 DNA densities and the percentages of 13C6-d-glucose in the substrate (Fig. 2b, R2 = 0.952), resulting from the link between the isotopic composition of the host and the produced virions. Since we observed a correlation between the 13C content of the growth substrate and the isotopic composition of both the cells and T4 DNA, we can conclude that there is a correlation between the isotopic composition of the host and the virions they produce, as expected, although the correlation coefficient was not directly measured.
Fig. 2
Buoyant density of T4 bacteriophage DNA measured in CsCl gradient. aT4 DNA density as a function of the proportion of 13C6-d-glucose employed in the d-glucose substrate for host growth. b T4 DNA density as a function of the theoretical density calculated according to the equation presented in the text [10]. The identity line is shown with red dots. The dashed line and equations shown on the plots are related to the linear regressions. The data originate from 2 distinct experimental series. In experimental series A, 0% and 100% 13C6-d-glucose proportions were tested, in biological triplicates. In experimental series B, 0%, 20%, 40%, 60%, and 80% 13C6-d-glucose proportions were tested, in biological triplicates. However, there was an experimental problem for one replicate of the 40% condition and one of the 60% condition, so that there are only duplicates shown in these cases
Observed T4 DNA Densities Are Greater than Expected from Empirical ModelsWe calculated the theoretical expected density for T4 DNA, by relying on a previously established empirical formula [11]:
$$\rho_ = 1.660 + \left( \right]} \right) + 0.036 \times \left[ ^C} \right],$$
where \(\left[G+C\right]\) is the GC content of the considered DNA (0.3530 for T4, NC_000866.4) and [13C] is its 13C content.
The obtained predicted densities were significantly lower than the measured ones (Fig. 2b). T4 DNA contains glucosylated hydroxymethylcytosine (HmC) instead of cytosine [12], affecting its buoyant density: it was detected as heavier than expected without DNA modification, in a previous study [13], with a value of 1.698 compared to 1.694 according to the above formula. In the present study, an average value of 1.701 (± 0.004 STD) was obtained for unlabeled T4 DNA, based on duplicates. For the fully labeled T4 DNA, the average density value was of 1.744, compared to an expected value of 1.731 according to the empirical model. Based on the linear regression from Fig. 2a, fully label T4 DNA has a density increase of 0.042 (compared to 0.036 in the above model). Models linking the GC content to the buoyant density would thus need to be adjusted for modified DNAs. Here, we do not suggest a new model, as it would require the analysis of DNA presenting the same modifications as T4, but with various GC contents.
Viral DNA SIP: Powerful Tool to Establish the Link Between Virus and Its HostSeveral experimental methods are already available to identify hosts of viruses, including digital droplet PCR [14], proximity ligation [15], epicPCR [16], or viral tagging [17]. To our knowledge, none of these methods are trivial to implement, each possessing distinct advantages and limitations. While some methods are targeted, such as those based on PCR, others require the cultivation of hosts, as is the case with viral tagging. Proximity ligation, conversely, is untargeted but necessitates meticulous design and data treatment to mitigate noise.
Experimental techniques utilizing stable isotopes, such as DNA Stable Isotope Probing (DNA SIP) or NanoSIMS, complement the aforementioned approaches in host–virus interaction studies, particularly as they relate to metabolic activity. This aspect is crucial for functional ecology, with these methods showing significant promise in elucidating the connections between viruses and major biogeochemical cycles. They are thus worth further developing and investigating. As mentioned in the introduction, DNA SIP has been successfully applied in viral ecology studies, using either cellular DNA [3, 4, 8] or total DNA [6]. We postulate that applying SIP to viral DNA separately, although challenging, would be complementary and would provide a different view on the viral diversity, with increased sensitivity and, likely, a better detection of purely virulent viruses. Compared to viral DNA, cellular DNA or total DNA is indeed expected to be enriched in temperate virus sequences, present in cellular genomes. Moreover, since cellular DNA is dominant in the total DNA, one can expect a lower sensitivity for detecting DNA from low abundant, purely virulent viruses, compared to an approach focusing on the DNA directly extracted from virions.
However, in viral DNA SIP that targets a specific metabolism, only a limited proportion of the microbial community is likely to be labeled, which poses a notable challenge. This technique requires a considerable amount of environmental viral DNA for analysis. Based on our experimental data, at least 400 ng of viral DNA from an environmental sample is required to reliably discriminate between the 12C and 13C buoyant density peaks. This requirement highlights the importance of careful experimental design and emphasizes the need for efficient viral DNA extraction and concentration techniques to meet the threshold for successful isotopic discrimination.
Comments (0)