
Remember me

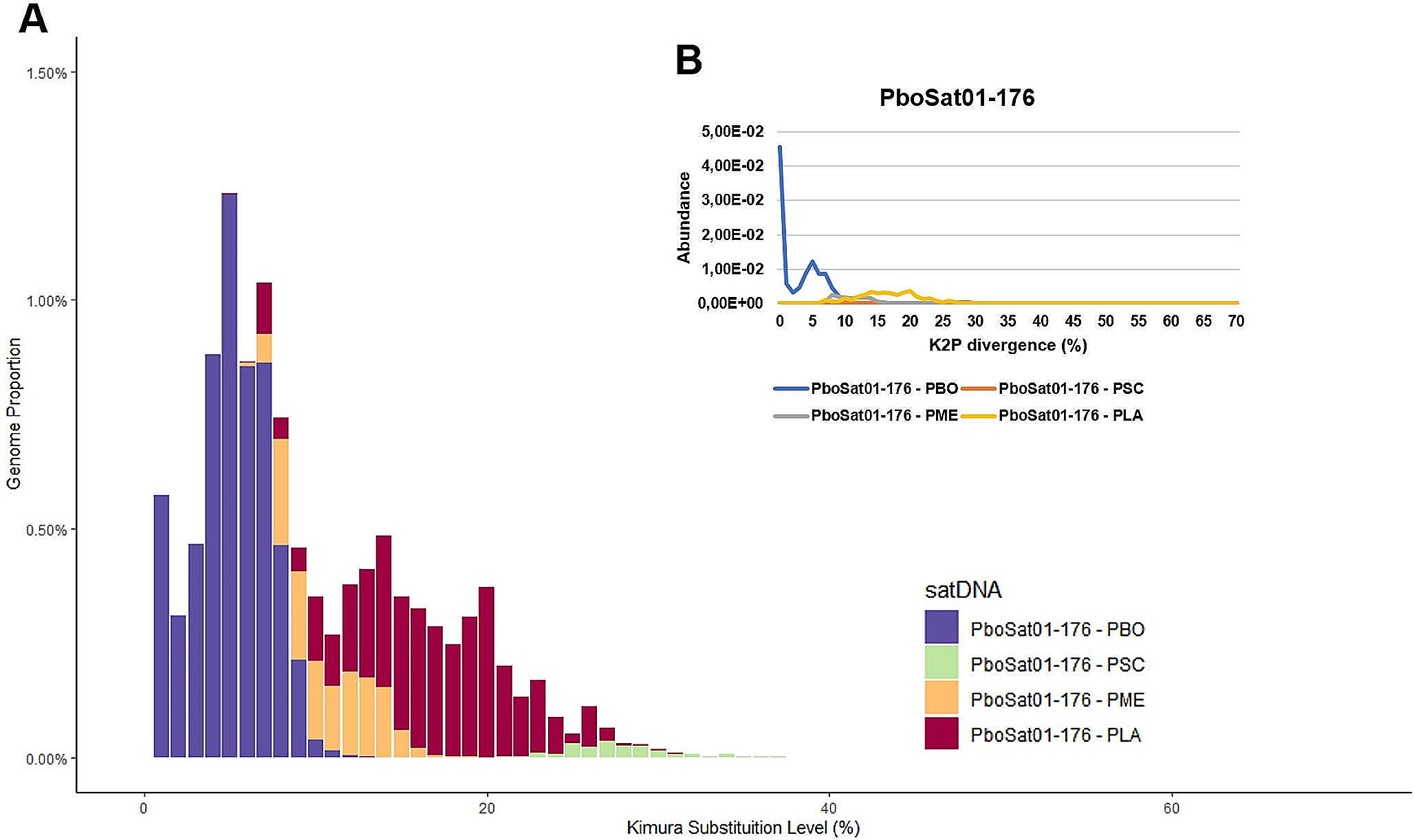






An even more striking manifestation of the non-randomness of COs/chiasmata is the nearly absolute rule that homologous-chromosome pairs almost always have at least one CO (e.g. no chiasmata, that is univalence, in only 4 among 25,120 pairs in C. brunneus spermatocytes, Jones 1987; review in Jones and Franklin 2006). This is true regardless of chromosome size (e.g. in the Zebra finch Taeniopygia guttata with 14 macro chromosomes and 64 micro chromosomes or the Japanese quail with 7 macro chromosomes and 31 micro chromosomes, each with at least one Mlh1 focus (Fig. 6A; Calderón and Pigozzi 2006). This feature is often referred to as the "obligatory crossover" because it reflects the fact that at least one CO per bivalent is required to satisfy the mechanical requirement for homolog connectedness at the first meiotic division. Thus, from the point of view of the meiotic process per se, this is probably the most important feature of CO non-randomness. In the standard meiotic program, occurrence of the obligatory CO is a specific, programmed effect. This is emphasized by the fact that, in the vast majority of organisms, the total number of COs per bivalent is very small, often two or a handful, and sometimes one and only one (review in Mercier et al. 2015); but the obligatory CO rule is nonetheless observed.
Fig. 6
Synaptonemal complex and recombination nodules. A MLH1 foci ( white) along pachytene chromosomes (red) in zebra finch (Taeniopygia guttata), 2n = 80 comprising 14 macro-chromosomes and 64 micro-chromosomes (Calderon and Pigozzi, 2006). B Diplotene chiasmata in desert locust Schistocerca gregaria with different sizes of chromosomes (Jones and Franklin 2006). C EM image of SC and EM-defined SC-associated nodule (arrow) that correspond to crossover recombination complexes, in D. melanogaster female. Note that the nodule does not penetrate the SC central region (Carpenter 2003). D EM spread of coaligned homolog axes linked by bridges (arrowhead) in spread preparations of Allium fistulosum (Albini and Jones 1987). E EM-defined SC-associated nodule (RN) is confirmed as a crossover site by immunogold colocalization of crossover factor Mlh1 (Lhuissier et al. 2007)
The lengths to which organisms go to ensure the obligatory CO is further manifested in the fact that it is observed: (i) irrespective of the total number of chromosomes (from two in the worm Parascaris univalens and in the Australian daisy to over 1400 in some adders-tongue ferns like Ophioglossum reticulatum); (ii) in species having a wide range in chromosome sizes like the grasshoppers Schistocerca gregaria (Fig. 6B) and C. brunneus (Jones 1986); (iii) in situations where the opportunity for COs is spatially restricted, e.g. when the chromosome is very short (e.g. Murakami et al. 2021; Calderón and Pigozzi 2006), when a majority of COs arise in less than a quarter of the genome due to the presence of large blocks of heterochromatin (e.g. Mercier et al. 2015) and for the XY sex chromosomes of mouse and human, where the available homologous region ("PAR") is tiny compared to total chromosome lengths (Kauppi et al. 2012). However, in few organisms, notably fission yeast and Aspergilus nidulans (which, notably, also lack CO interference; above), occurrence of at least one CO per bivalent is achieved by having a sufficiently large number of COs (~ 10–20 per bivalent in both cases) that, by chance, the probability of zero events is extremely low.
It is now known that occurrence of at least one (obligatory) CO depends on two factors. First, at least one pre-CO recombination interaction must be designated to mature along a CO-specific pathway. This effect is likely to be an intrinsic consequence of the same CO patterning process that gives rise to CO interference (Zhang et al. 2014a, c; Zickler and Kleckner 2023). Second, the CO maturation process must occur efficiently, an effect sometimes referred to as "CO assurance" to indicate that additional events are required after "CO designation" to "assure" that a CO product actually occurs (Shinohara et al. 2008).
Ultrastructure analysis: the synaptonemal complex and recombination nodules.The synaptonemal complexUltrastructural electron microscopic (EM) studies of meiosis commenced in the 1950s with remarkable and still not fully understood findings. First, Moses (1956) and Fawcett (1956) revealed the presence of a tripartite core-like structures called synaptonemal complexes (SCs), that run along the entire lengths of the homologs. SCs have further been found in most plants, animals, fungi, algae and protozoans that reproduce sexually (with the known exceptions of fission yeast, A. nidulans, D. melanogaster male and the protist Tetrahymena thermophila, review in Loidl 2021). The SC forms by linking the structural axes of two homologs by a close-packed array of transverse filaments which emanate towards one another from the two axes and with additional specific proteins located at the points where they converge (review in Page and Hawley 2004; Fraune et al. 2016; Zwettler et al. 2020). By EM, these features comprise the SC "central region" and its component "central element" which defines the midline of the central region along its length (Fig. 6C).
The SC was shown to normally form between homolog axes, as illustrated for example in reciprocal translocations, where chromosome segments are exchanged between two non-homologous chromosomes (review in von Wettstein et al. 1984). However, when no partner is available, SCs can assemble between nonhomologous axes in which the two axes are close together (e.g. in foldback chromosomes), between non-homologous chromosomes in haploid meiosis or between the three homologous chromosomes in triploid strains (examples in Rasmussen 1977a, b; Rasmussen et al. 1981; von Wettstein et al. 1984; Zickler and Kleckner 1999). Also, as observed in many species, SC components have a tendency to self-assemble outside of chromosomes, e.g. when the normal assembly process is defective, leading to stacks of SCs called polycomplexes (review in Hughes and Hawley 2020). Recent studies have now revealed that SC central region components are highly dynamic, suggesting that the structure meets the definition of a liquid crystal (Rog et al. 2017; Zhang et al. 2020).
Interestingly, the SC can also play a direct role during meiosis I segregation: in achiasmate meioses (like in silk-worm females), a “modified SC” remains between homologs and provides thus the connection required for their proper segregation at Anaphase I (Rasmussen 1977a, b, review in Zickler and Kleckner 1998). It was, however, shown recently that the so-called modified SC, is in fact formed by the fusion of two SC lateral components HOP1 and SYCP2 with a thick HOP1 band sandwiched by two layers of SYCP2. The authors propose to rename this structure “bivalent bridge” (Xiang et al. 2024).
EM analysis of 3D reconstructions and spread preparations of mammals, plants and fungi revealed that before SC formation, the axes of homologs are first coaligned at a distance of ~ 400 nm (e.g. Albini and Jones 1987; Anderson and Stack 1988; Zickler 1977; 2006). Such images revealed that the fundamental task of homolog recognition and juxtaposition precedes SC formation, with genetic studies further showing that both processes are dependent on recombination (e.g. Storlazzi et al. 2003; Dubois et al. 2019). Furthermore, coaligned axes are sometimes linked by EM-visible bridges that often exhibit nodular structure (Fig. 6D; Albini and Jones 1987; Moens et al. 2002; Anderson and Stack 1988; Anderson et al. 2001); and sometimes nodules also occur in association with individual homolog axes which are not visibly linked to any partner (Albini and Jones 1987; Anderson et al. 2001; Moens et al. 2002; 2007). Such images suggested that recombination complexes are associated with chromosome structures (axes/SCs) from very early (presynaptic) stages onward and that SC nucleation occurs at sites of recombination interactions via bridge structures (Dubois et al. 2019).
These suggestions were directly validated by: (i) fluorescence imaging of axes, bridges and recombination complexes both before, and at, the coalignment stage (e.g. Oliver-Bonet et al. 2007; Moens et al. 2007; Storlazzi et al. 2010; Dubois et al. 2019); (ii) EM documentation of SC nucleation at sites of recombination nodules (Zhang et al. 2014a); (iii) identification and molecular analysis of SC-nucleating bridges, which were shown to be “mini-axes” containing cohesin plus recombination components (Dubois et al. 2019) and (iv) that bridges mediate transfer of recombination complexes from on-axis to between axis/on-SC localization (Dubois et al. 2019; review in Zickler and Kleckner 2023).
Recombination nodulesUltrastructural analyses also revealed nodule-like structures associated with SCs (Fig. 6C). In a groundbreaking study of D. melanogaster oogenesis Carpenter (1975) argued that EM observed nodules at pachytene represent the sites of genetical COs because they exhibited the same pattern of obligation and interference that had been detected by genetic studies and called them “recombination nodules”. Other evidence for a correspondence between COs and SC-associated nodules was provided by the observation that in the plant Allium fistulosum nodules were proximally localized, in agreement with the distribution of chiasmata observed at later stages (Albini and Jones 1988). Many subsequent investigations identified similar nodules in several plant, animal and fungal species and showed that: (i) every SC had at least one nodule, (ii) that they corresponded to the number of chiasmata, (iii) that they displayed interference and, finally, that they were maintained at chiasma sites (with remnants of SC pieces) up to diakinesis (reviewed in von Wettstein et al. 1984; Zickler and Kleckner 1998; 1999). Also, EM analysis of recombination-defective mutants showed that the number and distribution of nodules parallels the number and distribution of genetically observed COs (Carpenter 1979; Zickler et al. 1992). Subsequent fluorescence imaging studies confirmed this correspondence to CO recombination machineries: recombination-associated proteins (notably MLH1/3, implicated directly in late stages of CO formation, and the E3 ligase Hei10) co-localize with such nodules at both EM and LM levels (e.g. Marcon and Moens 2003; Anderson et al. 2014).
These EM-visualized structures that correlated with COs later became known as "late recombination nodules" because observations made on earlier prophase I stages described another type of nodules, often smaller, much more numerous and already seen at the "zygotene" stage (Carpenter 1987; von Wettstein et al. 1984; Albini and Jones 1987; Bojko 1989; Anderson and Stack 2005). These were first termed "zygotene nodules" and, later, early recombination nodules. However, in some organisms (e.g. in the fungus Sordaria macrospora and tomato), nodules that correlate with COs and are morphologically distinct from the other, more numerous nodules, are already present at zygotene (Lhuissier et al. 2007; Zhang et al. 2014a). Immunogold staining in EM and recent light-fluorescence imaging studies showed that those early nodules correspond to the localization of recombination proteins like Rad51, RPA, Mer3 and Msh4/5, required for early stages of recombination (e.g. Moens et al. 2002; de Boer et al. 2006; Oliver-Bonet et al. 2007; Storlazzi et al. 2010; Yokoo et al. 2012) while late nodules correspond specifically to proteins involved in formation of crossovers (e.g. Mlh1, Fig. 6E; Lhuissier et al. 2007; Anderson et al. 2014).
It is now clear from many studies of meiosis that a large number of recombination interactions are initiated very early, among which only a few ultimately mature into COs. That is: "many are called but few are chosen" (to be COs). This relationship emerged originally from genetic studies of recombination in fungi which identified and analyzed non-Mendelian marker segregation, which is diagnostic of DNA/DNA interactions for recombination, as well as CO (Lindegren 1933; Zickler 1934; reviewed by Whitehouse 1982). Non-Mendelian segregations were found to be much more frequent than COs. Moreover, non-Mendelian segregation of a particular allele could be accompanied, or not, by crossing-over between markers at flanking positions, with the CO outcome usually being the rarer of the two (review in Whitehouse 1982). Put more specifically: many DNA/DNA interactions occur, among which a small subset is matured as COs (with accompanying interference and obligation) while the remainder are matured without crossing-over, i.e. as "noncrossovers" (review in Hunter 2015). The so-called "early nodules" may correspond to total recombination complexes and/or only to those that will not become COs and thus have been left behind, according to the situation. Presciently, Carpenter proposed that early nodules corresponded to noncrossovers and that the excess of such interactions were more abundant than COs because they were involved in homolog pairing (Carpenter 1987).
Further analysis of EM data revealed that, in Sordaria, SC nucleation sites are evenly spaced (in accord with observation from Allium images Albini and Jones 1987) and confirmed the implication that the classical interference process, which gives evenly-spaced COs along the chromosomes, actually acts more broadly to give a larger number of evenly-spaced SC nucleation sites, a subset of which are (interfering) CO sites (Zhang et al. 2014a). This and other findings point to the occurrence of a designation/interference process that operates at the structural level, with classical CO interference resulting from coupling of recombination complex status to that process (see discussion in Zickler and Kleckner 2023).
The bouquet and interlockings: two sides of the pairing coinEarly cytological studies identified two interesting chromosome configurations that occur contemporaneously during the period when homologs are undergoing pairing and synapsis. One such configuration is the "bouquet", in which all chromosomal telomeres are tightly clustered in a small region of the nuclear envelope and, in classical images, homologs are not obviously paired. First described and named by Eisen (1900) in salamander meiosis and further described in detail by Gelei (1921; Fig. 4B), this configuration has since been found in most studied organisms (review in von Wettstein et al. 1984; Scherthan 2001; Zickler 2006; Zickler and Kleckner 1998; 2016). A second interesting configuration is whole chromosome "interlocking", in which one chromosome, or a pair of homologs, is trapped within an open, unpaired region of another chromosome pair (examples in von Wettstein et al. 1984; Rasmussen 1986). Interlocked configurations emerge during the pairing period but are then mostly actively resolved by the end of prophase, with only regular bivalents seen by the end of pachytene (e.g. Storlazzi et al. 2010).
The bouquet configuration was originally proposed to occur prior and prerequisite to pairing of homologous chromosomes as a way of topologically simplifying the homology search process (e.g. Gelei 1921; Scherthan 2001). Recent studies reveal that the bouquet configuration is the result of active telomere-led movements driven by the cytoskeleton and that it can be quite transient (review in Link and Jantsch 2019). Early studies also identified global whole-nucleus rotations (Parvinen and Soderstrom 1976), now also understood to be mediated by cytoskeletal forces. Diverse lines of evidence suggest that while the bouquet configuration and or cytoskeleton-mediated movements may promote juxtaposition of homologs, chromosome motions also appear to provide a stringency factor which has the potential to eliminate both unwanted pairwise associations and whole chromosome interlockings (e.g. review in Link and Jantsch 2019; Klutstein and Cooper 2014; Zickler and Kleckner 2016 and references therein).
Importantly, also, elimination of interlocks requires not only regularization of whole chromosome topological relationships but resolution of constraining DNA interactions. This is revealed by the finding, in Sordaria, that interlock resolution requires Mlh1 (Storlazzi et al. 2010). Mlh1 is a subunit of a structure-specific nuclease that not only plays a central role in crossover formation (Hunter
Comments (0)