
記住我









Activation of a conditionally functional dicentric chromosome in which the two centromeres are homologous results in the generation of monocentric derivatives. Homology-based repair pathways include reciprocal crossing over (RCO, Fig. 1A) and single-strand annealing (SSA, Fig. 1B) when the centromeres are oriented as direct repeats (Brock, et al. 1994). Repair efficiency can be estimated from 60 to 70% of cells that survive dicentric chromosome activation (Cook, et al. 2021). High cellular viability together with repair through centromere homology leads to the question of whether the centromere is a preferred site of recombination.
Fig. 1
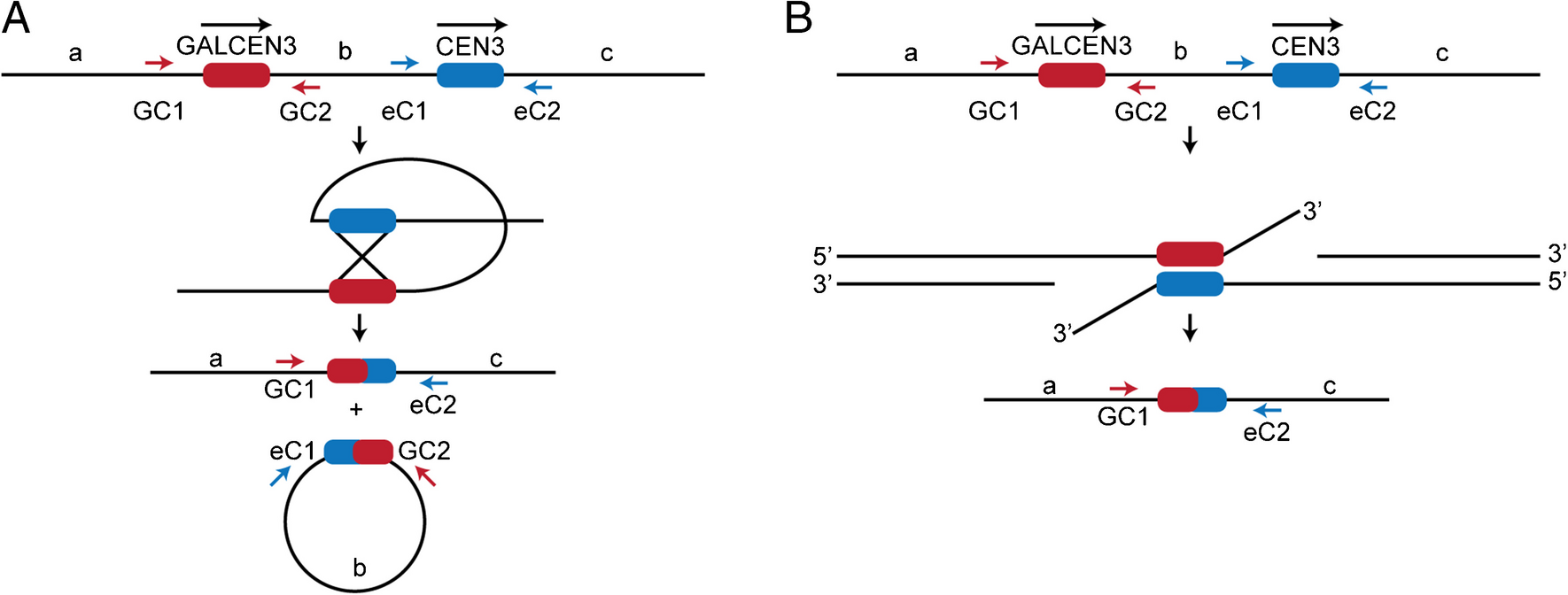
Centromere Homology Repair Pathways. A Schematic of the Reciprocal Crossover Event (RCO) that is the dominant repair pathway in dicentric chromosomes with 46.3 kb between the two centromeres. The two centromeres (GALCEN3 and CEN3) are in direct orientation. Primers used to amplify parental and recombinant products are marked. Recombinant centromeres arise in the repair products; GC1-eC2 and eC1-GC2 are a result of the two RCO events. B Schematic of the Single-Strand Annealing Event (SSA). This event yields a recombinant centromere that can be identified with GC1-eC2 primers
To examine the kinetics of repair, we quantitated the appearance of recombinant products following activation of the Gal1-regulated centromere (GALCEN3) at varying distances from the endogenous CEN3 (Fig. 2). The parental centromeres, CEN3 and GALCEN3, were detected through PCR with eC1-eC2 (endogenous centromere) and GC1-GC2 (Gal1-regulated centromere) (Fig. S1) and the recombinant products with eC1-GC2 (endogenous Cen to GalCen) and GC1-eC2 primers (GalCen to endogenous Cen) (Fig. 2). Fluorimetry quantitation of recombinant products shown in Fig. 2 reveals these products to be far from stoichiometric (note scale bars on y-axis in Fig. 2). Both recombinant products increase over time from 2 to 24 h (Fig. 2). Time points from 2 to 6 h are reflective of initial events following dicentric chromosome activation, while accumulation at later time points (24–72 h) reflect selective advantages in growth or cell cycle progression. There is a 2-fold variation in the early time points for the GC1-eC2 and eC1-GC2 products (at 6 h), but there is no relation to inter-centromere distance (18.2 kb > 12.3 and 9.8 > 46.3 > 6.5) (Fig. S2).
Fig. 2
Recombinant products after dicentric chromosome activation. A Fluorimetry quantitation of recombinant PCR product GC1-eC2 in strains with GALCEN3 inserted 6.5, 9.8, 12.3, 18.2, and 46.3 kb away from the endogenous CEN3, after switching carbon source from galactose to glucose (GALCEN3 activation) and growing for indicated times. From 0 to 6 h, the kinetics of GC1-eC2 product generation are similar for all strains (see Fig. S2). The GC1-eC2 product is the result of RCO between the two centromeres (Fig. 1A) and the non-reciprocal SSA event (Fig. 1B). The time courses represent data from three independent cultures. N=9 for each data point, error bars are the standard error of the mean. B Fluorimetry quantitation of PCR product eC1-GC2. Note the difference in y-axis scaling compared to A, eC1-GC2 is not stoichiometric with GC1-eC2. The 46.3 kb dicentric is the only strain with markedly elevated eC1-GC2 product after 72 h due to selection for the RCO event at this CEN-CEN distance. The time courses represent data from three independent cultures. N = 9 for each data point, error bars are the standard error of the mean
The eC1-GC2 product is significantly reduced relative to GC1-eC2 (Fig. 2). In addition, the eC1-GC2 product is most evident in the 46.3 kb dicentric (from 6 to 72 h, Fig. 2B), versus in the pericentric dicentrics (18.2, 12.3, 9.8, and 6.5). The marked distance-dependent relationship in the eC1-GC2 product is consistent with the preferential generation of the ring and rod monocentric derivatives found in the 46.3 kb dicentric. Reciprocal crossover (RCO) between GALCEN3 and CEN3 is the preferred repair event in dicentric chromosomes with 46.3 kb between the two centromeres due to the selection for an essential gene (NFS1, 20 kb from CEN3) between the two centromeres (Cook, et al. 2021).
Distinguishing homologous recombination pathways in dicentric chromosome repairThe disparity in stoichiometry between the recombinant products is indicative of the multiple pathways to generate monocentric derivative chromosomes. To investigate the preferred DNA repair, we used quantitative analysis of the PCR products in strains with the two centromeres oriented as direct repeats (shown in Fig. 1) or inverted repeats (inverse of GALCEN3 in Fig. 1). Quantitative analysis was performed at the 48-h time point to ensure ample time to observe the recombinant products, but prior to bias due to growth selection. The GC1-eC2 recombinant product is the dominant event (~10-fold) in all dicentric locations where the two centromeres are in the direct orientation (dark blue column, direct orientation, Fig. 3). The presence of the GC1-eC2 product is predicted from a double-strand break between the two centromeres, followed by resection and single-strand reannealing of the 340 bp shared between the two centromeres (see Fig. 1B). The GC1-eC2 product would also arise following break-induced repair via the invasion of one centromere into the second, followed by DNA synthesis (break-induced repair, BIR). In a BIR event, a break at GALCEN3 followed by resection is followed by a single-strand invasion of the GALCEN3 into the endogenous centromere, where the invaded strand serves as the template for DNA synthesis. Either of these non-reciprocal pathways leads to stable monocentric derivatives lacking intervening DNA, the length of which depends on the distance between the two centromeres.
Fig. 3
Stoichiometry of recombinant products as a function of position and orientation of the conditional centromere. Fluorimetry quantitation of GC1-eC2 and eC1-GC2 recombinant products after 48 h on glucose in strains with GALCEN3 inserted 6.5, 9.8, 12.3, 18.2, and 46.3 kb away from the endogenous CEN3, in both the direct orientation (Fig. 1A) and the inverted orientation, where GALCEN3 is flipped relative to the endogenous centromere. In dicentrics with centromeres in the direct orientation, the GC1-eC2 product (the results of RCO and non-reciprocal SSA) dominates the eC1-GC2 product, which is only produced through RCO. In strains with inverted oriented centromeres, SSA events lead to iso-dicentrics that rearrange in subsequent divisions (see text) and therefore do not accumulate in the population. The time courses represent data from three independent cultures. N = 9 for each data point except no product control where n = 3, error bars are standard error of the mean
Recombination between centromeres oriented as inverted repeats does not generate stable monocentric derivativesThe recombinant products in strains with inverted-orientation centromeres (GALCEN3 inverted relative to CEN3) are drastically different in yield relative to the direct orientation (Fig. 3). The major difference is the reduction of the GALCEN3-CEN3 (GC1-eC2) product (dark blue column, Fig. 3). Reciprocal cross-over between inverted centromere repeats generate a dicentric derivative chromosome with an inversion of DNA between the two centromeres, or broken fragments with one telomere and one broken end (the right and left chromosomal arms, respectively). SSA between centromeres in the inverted orientation results in iso-dicentric chromosomes through intra-chromosomal rearrangement or truncated chromosomes due to fold-back structures (Ramakrishnan, et al. 2018). In either case, stable monocentric derivatives are not generated. Rearrangement chromosomes containing the GC1-eC2 product from strains with inverted centromeres will continually rearrange until events arise that generate the monocentric derivative. The differences in the yield of recombinant products in the direct vs inverted dicentric strains can be accounted for by the inability of SSA or BIR events to yield stable monocentric derivatives when the regions of centromere homology are inverted with respect to one another.
SSA requires the action of single-strand endonuclease RAD1/RAD10 to process the non-homologous tails flanking the region of homology (Fig. 1B) but does not require the product of RAD51, which is required for strand invasion. To investigate the contribution of SSA to dicentric repair, we examined the kinetics of recombinant products in the dicentric chromosomes with centromeres in the direct orientation and separated by 46.3 kb in rad1Δ and rad51Δ mutants (Fig. 4A). The yield of GC1-eC2 product is dependent on Rad1 and independent of Rad51 (Fig. 4A), consistent with a non-reciprocal single-strand annealing (SSA) event dominating the repair kinetics when centromeres are in the direct orientation (Fig. 3) (Brock, et al. 1994); (Thrower, et al. 2003).
Fig. 4
The GC1-eC2 Recombinant product is dependent upon Rad1 but not Rad51. A Fluorimetry quantitation of the recombinant PCR product GC1-eC2 in the 46.3 kb dicentric, WT, rad51Δ, and rad1Δ. WT data is from Fig. 2. The GC1-eC2 recombinant product is reduced in rad1Δ. N = 9 for each data point. Error bars are standard errors of the mean. Cell viability is reduced in the 46.3 kb rad51Δ strain, (45.1 ± 2.0%) compared to WT (61.0 ± 2.1%) (Cook et al., 2021), student’s t-test p-value < 0.01. Cell viability is not reduced in the 46.3 kb rad1Δ (54.5 ± 1.9%) compared to WT (61.0 ± 2.1%) (Cook et al., 2021), student’s t-test p-value > 0.01. Error is the standard error of the mean, n = 9 for each viability assay. B Fluorimetry quantitation of the recombinant PCR product GC1-eC2 in the 46.3 kb and 9.8 kb dicentrics, WT, and rad51Δ. WT data is from Fig. 2. GC1-eC2 is unaffected in rad51Δ. N =9 for each data point. Error bars are standard errors of the mean. The 9.8kb rad51Δ dicentric strain exhibits reduced viability (48.3 ± 1.1%) compared to WT (62.4 ± 1.2%) (Cook et al., 2021), student’s t-test p-value < 0.01. Error is the standard error of the mean, n = 9 for each viability assay
The predominance of non-reciprocal homologous recombination in strains with the direct orientation provides the opportunity to estimate the site of breakage between the two centromeres. The requisite for SSA is the processing of DNA through 5′-3′ resection until single strands from each centromere become available for annealing with one another. The kinetics of SSA along dicentric chromosomes with very different centromere distances will distinguish whether breaks occur at random between the two centromeres, or whether there might be preferred break sites. These experiments were performed in the absence of Rad51 to rule out breaks within the centromere that could generate a distance-independent repair product via BIR. We examined the kinetics of the GC1-eC2 recombinant product in rad51Δ mutants in dicentric chromosomes with centromeres separated by 9.8 kb or 46.3 kb, respectively (Fig. 4B). The kinetics are comparable in the two strains from 0 to 6 h (Fig. 4B) and within the range of variation observed in wild-type strains in which there is no progression in the amount of product vs inter-centromere distance (Fig. 2). Thus, SSA is the predominant homology-based repair pathway responsible for the conversion of the dicentric to a monocentric deletion derivative (Figs. 3 and 4). The similarity in the kinetics of the GC1-eC2 products between centromeres whose distances range from 6.5 to 46.3 kb points toward the possibility that the initiating break sites are not randomly distributed between the two centromeres, rather they are biased toward the centromeres as reported by Song et al. (2013).
Kinetics of single-strand annealing is distance-independentQuantitative analysis of PCR products reveals that the kinetics of accumulation of the GC1-eC2 SSA product from 2 to 6 h on glucose is largely independent of the distance between the centromeres (Figs. 2 and S2). Considering the 7-fold change in distance (6.5 to 46.3 kb) and the rate of resection (3–4 kb/hr) (Yan, et al. 2019), we expected to see a difference in kinetics from 1 to 5 h (1 h for the 6.5 kb to 5 h for the 46.3 kb dicentric chromosome, respectively). Instead, the kinetics are comparable from 0 to 6 h (Figs. 2 and S2).
These data point to an initiating double-strand break or single-strand nick for DNA repair in the region of centromere homology rather than random breakage between the two centromeres. Prior studies have also pointed to the rapid appearance of the SSA repair product following the activation of the dicentric chromosome. Brock and Bloom (1994) observed the SSA product at 2.5–5 h following the growth of glucose. In contrast, the appearance of an SSA event via direct HIS4 repeats was not apparent until 12–24 h (Brock, et al. 1994). In these dicentric chromosomes, a 1.56-kb region of the 5′ end of the HIS4 gene is duplicated, with the repeats on either side of the GALCEN3 sequence. Activation of the dicentric stimulates non-reciprocal exchange between the HIS4 repeats, and the resulting product was observed at 12–24 h (Brock, et al. 1994). To confirm and extend these results, strains containing the dicentric chromosome with centromeres in the direct orientation and flanked by HIS4 repeats (Fig. 5B) were transferred to glucose, and the recombinant products were identified by PCR over time. As previously found, the centromere recombinant product (GC1-eC2) was observed by 4–6 h growth on glucose. In contrast, the HIS4 rearrangement was observed at 24 h (Fig. 5A). The centromeres are 46.3 kb from one another and share 340 bp of homology, while HIS4 repeats are 5.5 kb from one another and share 1.56 kb of homology. These data indicate that random breaks between repeats are unlikely to be initiating events responsible for centromere exchange. In contrast, if breaks are generated within the centromere, the prediction would be that a few hundred bp of resection will expose the two single-stranded centromeres for annealing, leading to rates of SSA independent of centromere–centromere distance. Alternatively, a DSB in one centromere may initiate a non-reciprocal recombination event with the unbroken centromere such as those observed in break-induced replication (Liu, et al. 2022).
Fig. 5
Kinetics of HIS4 vs CEN3 recombination. A Measurements of band intensity of recombinant PCR products in two different 46.3 kb dicentric strains. Strains are diagrammed in (B). Recombination between the two HIS4 repeats is kinetically delayed compared to recombination between CEN3 and GALCEN as measured by the GC1-eC2 product (blue line). The RCO product, eC1-GC2 (orange line), appears at the same time at the HIS4 rearrangement (green line) 24 h after activation, whereas GC1-eC2 appears at 4–6 h. There is a temporal bias toward CEN-CEN recombination that is not explained by homology or distance, since the HIS4 strain has more homology and shorter distance between repeats, which should facilitate homologous recombination at a faster rate than that between two distant centromeres. This bias could be the result of breaks within the centromere as opposed to random breaks between the two repeats. N = 2 for each data point. B Schematics of the two different dicentric chromosome constructs used in this assay. One strain has GALCEN3 (red) positioned 46.3 kb away from the endogenous CEN3 (blue) in the direct orientation. These identical centromeres only have 340 bp of homology to facilitate homologous recombination. The other strain has an identical dicentric construction but has a duplication of 1.5 kb of the HIS4 gene, with the repeat on the other side of the GALCEN3 sequence. The two HIS4 repeats are separated by 5.5 kb. Upon dicentric activation, recombination occurs via the HIS4 repeats
Single-nucleotide polymorphisms in the centromeres reveal reciprocal crossover (RCO) events following dicentric chromosome activationThe initiating event for homologous recombination requires the invasion of a single-strand of DNA into a site of homology. The extent of homology between the two centromeres in the dicentric chromosome is 340 bp (Fig. 6A). Assuming a random distribution of breaks between the two centromeres, we would expect breaks at the centromere proper to represent less than 1% of the total inter-centromeric length (340/46,300 = 0.7%) in the 46.3-kb dicentric strain. The high viability (60–70%) of cells following dicentric chromosome activation (Brock, et al. 1994; Cook, et al. 2021) and the high fraction of survivors containing two reciprocal recombinant products (79% of survivors of the 46.3-kb dicentric contain GC1-eC2 and eC1-GC2 products) (Fig. 6B) reveals a discrepancy between the purported random distribution of breaks and the distribution of recombinant products in the survivors.
Fig. 6
Centromere SNPs cause a shift in recombinant products. A Diagram of the CEN3 sequence derived from s288C with single-nucleotide polymorphisms marked in light blue, at positions 33, 44-45, 74, 76, 87, 131, 133, 144, 164, 299, and 323. The divergent CEN3 sequence is derived from YJM789 and contains 12 SNPs, 7 of which are within the 117-bp core centromere. The CEN3 and flanking bases shown here are the 340-bp that are homologous between CEN3 and GALCEN3. These SNPs can be used to analyze recombinant products on a granular level. B Recombinant endpoint analysis of a strain containing the s288C-derived GALCEN3 at 46.3 kb and the YJM789-derived CEN3 at the endogenous position (1214-9). Single colonies were analyzed by PCR for CEN3 (eC1-eC2), GALCEN3 (GC1-GC2), and the two recombinant products (GC1-eC2 and eC1-GC2). HR was defined as both recombinant products present and no parental products. SSA was defined as GC1-eC2 only. Unrearranged was both parental products and no recombinant products. End-joining was characterized by a deletion in one of the two centromeres and no other recombinant products. Aneuploid events contained a recombinant product and one or both unrearranged parental products. The pattern of repair is dramatically shifted from 79% homologous recombination in the WT to 69% end-joining. A small fraction (7%) are HR events, with the remainder (24%) in the “unrearranged” category. N = 24 for WT (no SNPs) (Cook et al., 2021) and n = 59 for 1214-9 (12 SNPs). The glu/gal viability for 1214-9 is 55.9 ± 1.3% (student’s t-test compared to WT, p-value > 0.01). Error is the standard error of the mean, n = 9 for viability assay
To directly test the hypothesis that recombination initiated within the centromere triggers the RCO event, we introduced single-nucleotide polymorphisms (SNPs) into one of the two centromeres (Fig. 6A). A centromere from yeast strain YJM789 (Wei, et al. 2007) was introduced in place of CEN3 at its endogenous location in chromosome III (114,300 bp) in a strain derived from s288c. The second centromere (GALCEN3 68,000 at HIS4) was derived from s288c. The sequences of the two centromeres are shown in Fig. 6A. There are 12 SNPs within the 340-bp repeats, (33 bp, A/G; 44–45 bp, TG/AA; 74 bp, G/A; 76 bp, T/A; 87 bp, A/T; 131 bp, T/C; 133 bp, A/G; 144 bp, T/G; 164 bp, A/T; 299 bp, T/C; 323 bp, A/G). The 117-bp CEN3 spans 64–180 (inclusive) and has seven of the 12 SNPs.
Strains containing SNPs between the two CEN3s on chromosome III exhibited viability indistinguishable from strains with identical CEN3s (55.9 ± 1.3% viability glucose/galactose (student’s t-test compared to WT, p-value > 0.01). In contrast, the distribution of products in the surviving colonies was shifted between the two strains. With a dicentric strain in which the GALCEN3 is identical to the native CEN3 and the two centromeric sequences are located 46.3 kb apart (DCY1214.1), most (79%) of the survivors in cells grown in glucose contain reciprocal recombinants, one linear chromosome with a deletion and a circular derivative with the deleted sequences (WT, Figs. 1A and 6B (Cook, et al. 2021). In strains with 12-centromere SNPs (strains DCY1214-9 and DCY1214-21), the surviving population is comprised of 69% end-joining events in which one of the two centromeres has been deleted and 24% events in which the two centromeres remain in the parental configuration (unrearranged, Fig. 6B). The reduction in the rate of crossovers in the strain in which the centromeres have 12 SNPs (resulting in 97% identity) is not unexpected. Datta et al. (1997) showed that repeats that were 99% identical had a rate of homologous recombination that was about 10-fold less than repeats that were 100% identical. This reduction is caused by the action of the DNA mismatch repair system reversing heteroduplexes that have mismatches. About 7% of the events in the survivors of the activation of the dicentric chromosomes with heteroallelic centromeres contained the reciprocal crossover products (Fig. 6B).
The class of unrearranged centromeres likely reflects a different mechanism. There are several Ty elements on chromosome III (at 1kb, 84 kb, 149 kb, and 169 kb). An alternative pathway toward the generation of monocentric derivative chromosomes is recombination events between Ty1 elements that flank the centromere at positions 84 kb and 149 kb. In cells with centromeres in the inverted orientation (Hill, et al. 1989), or cells with non-homologous centromeres (Surosky, et al. 1985) in the same chromosome, the major products were a linear 65-kb monocentric deletion derivative and the reciprocal 65-kb ring chromosome, each with one of the unrearranged parental centromeres (Hill, et al. 1989; Surosky, et al. 1985). It is possible that the events labeled “unrearranged” in the strain with the heterozygous SNPs contain these Ty rearrangements. The introduction of SNPs between the two centromeres thus reduces the rate of CEN–CEN recombination and increases the frequency of other types of events.
To assess the molecular nature of recombination events between the two centromeres, we sequenced centromeres from cells harboring the RCO event. Single colonies were isolated as detailed in the “Materials and methods” section, and RCO cells were identified as individuals with only centromere recombinant products (eC1-GC2 and GC1-eC2 products, Figs. 1A and 6B, HR) and no parental products or end-joining events. Products from 19 single-colony isolates were sequenced to determine the recipient and donor strands and the extent of gene conversion.
A reciprocal recombination event can result from a crossover anywhere within the 340-bp duplicated region containing the centromeres. Although from Fig. 6B, one might expect that sequencing of the linear and circular products would reveal two recombinant junctions, we can only detect such junctions if the recombination event occurs between heterozygous SNPs. Thus, any event that occurs between the start of the homology and the SNP at position 33 bp or between the SNPs at position 323 bp and the end of the homology will not result in a detectable recombinant junction. In addition, many previous studies in yeasts and other organisms (Symington, et al. 2014) have shown that reciprocal crossovers are usually associated with a non-reciprocal exchange of information (gene conversion) located adjacent to the DSB that initiates the event.
Examples of the patterns of SNPs from the two different reciprocal recombinants are shown in Fig. 7. We divided the 19 analyzed recombinants into the following four classes: Class I (Fig. 7A 8 of 20), one chromosome with the parental arrangement of SNPs and one chromosome with a recombinant product; Class II (Fig. 7B 2 of 20), two chromosomes with centromeres with the parental (either both from one parent or one from each parent) configuration of SNPs; Class III (Fig. 7C, 5 of 20), both chromosomes with recombinant configurations of SNPs; and Class IV (Fig. 7D, 4 of 20), complex events in which one chromosome has two or more recombination breakpoints. In addition to the events depicted in Fig. 7, all of the sequenced events are shown in Fig. S3. For the recombinant products that have SNPs derived from only one of the centromeres, we infer that the breakpoint on this chromosome occurred in the regions of the duplication that did not contain heterozygous SNPs (proximal to the SNP at position 33 bp or distal to the SNP at position 323 bp) since all of the recombinant chromosomes had the PCR products expected for reciprocal exchanges (Fig. 6B). Seventeen of the 19 isolates had at least one detectable recombination breakpoint between polymorphisms. Of the 30 transitions between YJM789- and s288c-associated SNPs, 14 were within the centromeres, 11 were ambiguous (transition between a centromeric SNP and a flanking SNP), and 5 were unambiguously in the regions flanking the centromeres.
Fig. 7
Four classes of reciprocal recombinants resulting from the resolution of a dicentric. Each pair of lines represents independent reciprocal recombinant products with the linear product (line plus dot) on the top and the circular product (circle plus dot) on the bottom. The length of each map is 340 bp which is the amount of DNA duplicated in generation of the dicentric. The contributions of the s288c and YJM789 sequences are shown in red and blue, respectively. Vertical black lines inside the rectangles indicate the positions of heterozygous SNPs. The centromeric sequences are 117 bp of the 340 bp repeat (Fig. 6). The SNPs within the centromeres at the boundaries are shown with black arrows and the SNPs at the boundaries of the flanking sequences are shown as double-headed arrows. Maps of all of the sequenced recombinants are in Fig. S3
Based on previous recombination models (Symington, et al. 2014), in Fig. 8, we show the DNA interactions likely to generate the four classes of products illustrated in Fig. 7. We assume that the recombination events are initiated by a DSB, although nick-initiated recombination cannot be ruled out. The steps for all of the events are similar. Following cleavage of the DNA, the broken ends are degraded 5′ to 3′. One end invades the unbroken centromere, forming a heteroduplex. The invaded 3′-end is used as a primer sequence for DNA synthesis resulting in displacement of the paired DNA strand from the intact duplex. The displaced strand then pairs with the other broken end (second-end capture), resulting in regions of heteroduplex flanked by two junctions. If these junctions are cleaved in different planes (as shown in Fig. 8), a reciprocal crossover will be formed. If the heteroduplexes include SNPs, mismatched bases will occur (shown by yellow highlighting). The mismatch repair system will remove one of the mismatched bases. The 4 classes identified differ in several ways: (1) whether the s288c- or YJM789-derived centromere receives the initiating DSB, (2) the extent of processing of the broken ends, (3) the amount of DNA synthesized by the invading end, and (4) whether a mismatch is repaired to generate the s288c or the YJM789 SNP allele. In addition to the interpretive diagrams of four events shown in Fig. 8, we show similar diagrams for classes and sub-classes of reciprocal recombinants in Figs. S4–S7.
Fig. 8
Mechanisms for generating the recombinant classes (classes I-A, II-A, III-A, and IV-A). DNA strands are represented as black lines with the arrows showing the 3′ end of the strand. The red and blue circles represent s288c- and YJM789-derived SNPs, respectively. Following DSB formation, the strands are resected 5′ to 3′. One broken end invades the unbroken homolog, and DNA synthesis is primed from the 3′ invading end (shown by dotted lines). This synthesis results in the displacement of a DNA strand, allowing the displaced strand to pair with the other broken end. The resulting intermediate has two connecting Holliday junctions (HJ). If these junctions are cut by resolvases as shown by the triangles, the intermediate is resolved as a reciprocal crossover. Mismatched bases resulting from heteroduplex formation are highlighted by yellow boxes. Small arrows within the yellow boxes show the direction of mismatch repair (MMR). The red letters “A” and “B” and the blue letters “C and “D” represent the DNA sequences that flank the SNPs. Following the crossover, these flanking sequences are in the recombinant arrangement
This analysis leads to several interesting conclusions. First, the recombination events are initiated on the two centromeres of the dicentric chromosome with equal frequency. As shown in Fig. 8, consistent with many studies of recombination (Symington, et al. 2014), the sequence that has the initiating DSB is the recipient of information in the resulting gene conversion event. For example, as shown in Fig. 8 (I-A), when the s288c-derived centromere is broken, the resulting conversion event duplicates sequences derived from the YJM789 centromere. Of the 18 reciprocal events associated with gene conversion, 10 were initiated on the s288c GALCEN3, and 8 were initiated on the YJM789 CEN3. Second, the observation that, for most of the reciprocal recombinants, one of the interacting sequences contains exclusively s288c SNPs or YJM789 SNPs suggests that one of the junctions connecting the homologous sequences is resolved at the end of the homology (for example, Fig. 8 (I-A, II-A, and 8IV-A but not III-A). Lastly, although most heteroduplexes with more than one mismatch are repaired in the same direction (for example, Fig. 8 (II-A)), “patchy” repair is also observed (Fig. 8 (IV-A)).
Regulation of RCO productsDepending on the choice of which strands are cut to resolve Holliday junctions, homologous recombination is predicted to give rise to CO or NCO events. In the case of dicentric chromosomes, CO events lead to the generation of linear and circular monocentric derivatives (Cook, et al.
留言 (0)