
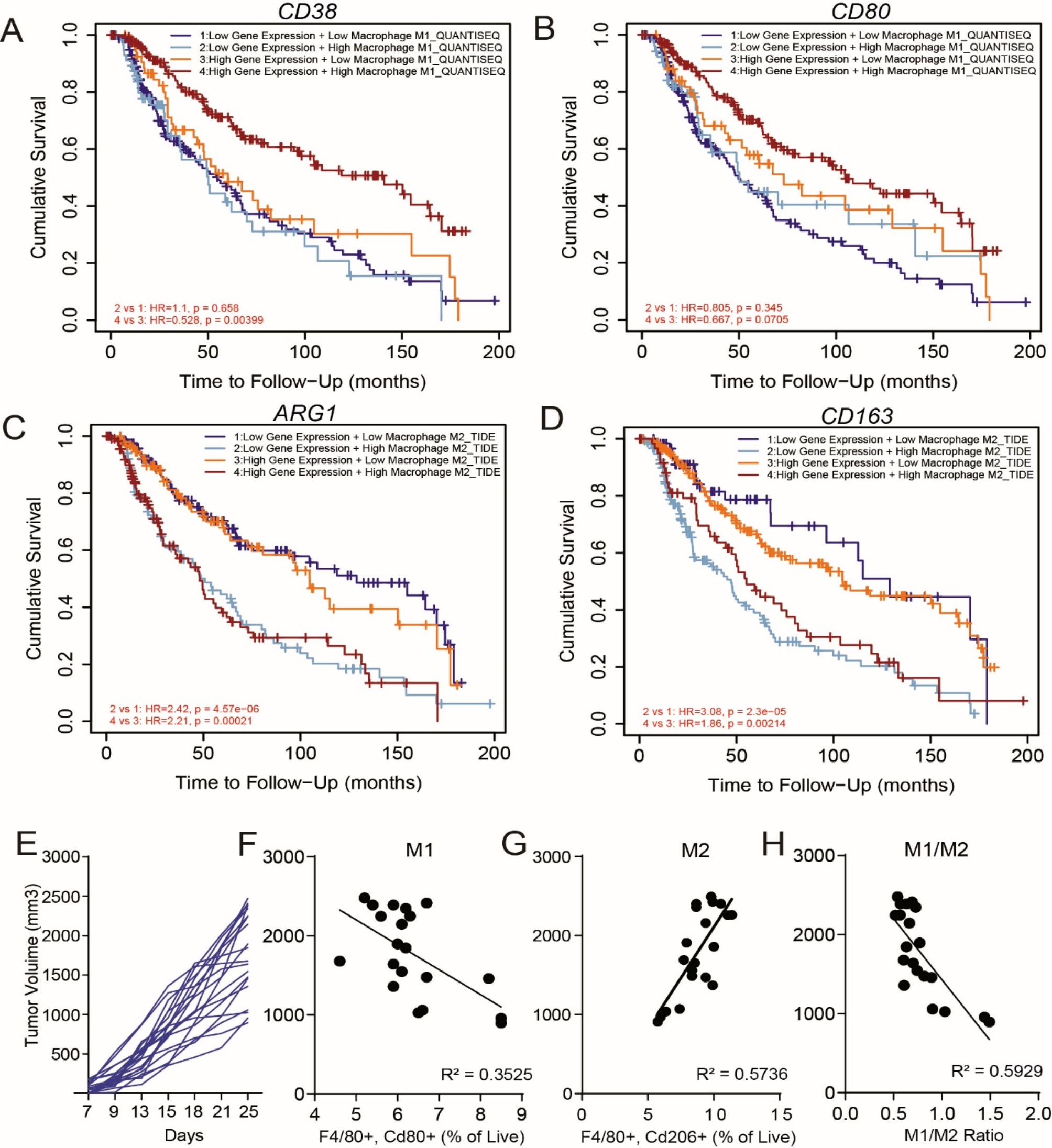
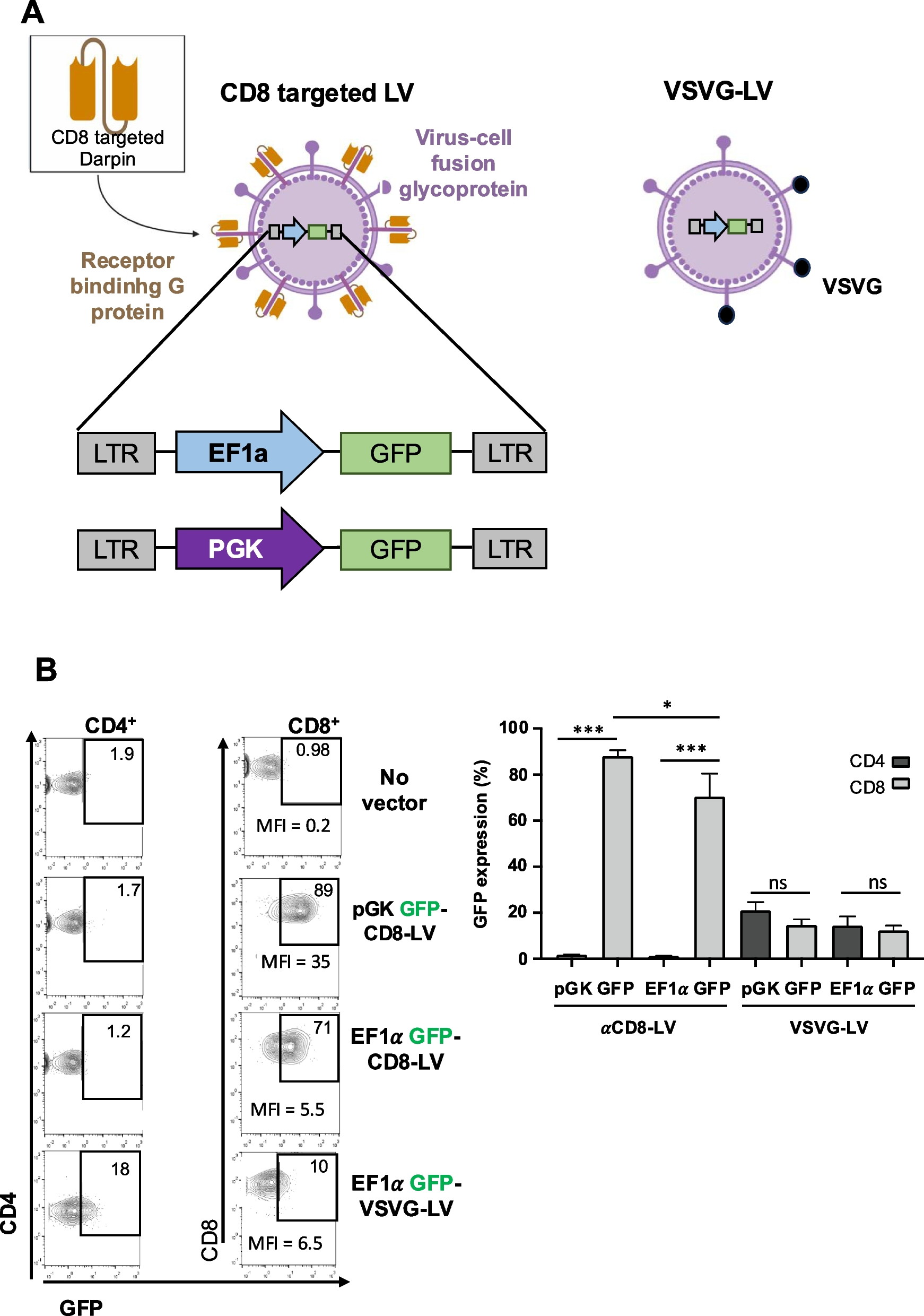
Remember me




Using a circRNA microarray, we identified 596 upregulated and 453 downregulated circRNAs in five CRC tissues compared with adjacent normal tissues (Fig. 1A). A clustered heat map revealed the differentially expressed top 20 circRNAs (Fig. 1B). Among these circRNAs, hsa_circ_0025506 (circGPRC5A) showed the greatest differential upregulation. CircGPRC5A is derived from exon 2 of GPRC5A and is located on chr12:13,061,176–13062105. To verify the closed-loop structure of circGPRC5A, we confirmed the presence of a splice junction in the PCR product of circGPRC5A by using Sanger sequencing (Fig. 1C). To further confirm the characteristics of circGPRC5A, we amplified circGPRC5A using divergent primers from cDNA, and not from gDNA, of CRC cell lines (HCT8 and HT29) (Fig. 1D, E). RNase R experiments demonstrated that the level of GPRC5A mRNA was reduced compared to levels of circGPRC5A (Fig. 1F, G). In addition, qRT-PCR demonstrated that the relative circGPRC5A levels were significantly higher in random hexamers than in oligo dT 18 primers (Fig. 1H, I). On the other hand, the relative circGPRC5 RNA levels were more stable than GPRC5A mRNA levels after actinomycin D treatment of HCT8 and HT29 cells (Fig. 1J, K). In conclusion, these findings indicate that circGPRC5A has a closed-loop structure.
Fig. 1
Human circRNA microarray analysis was used to characterize circGPRC5A in CRC. A Volcano plot of human circRNA microarray demonstrating upregulated or downregulated circRNAs in five CRC and paired normal tissues. B Human circRNA microarray cluster heat map displaying the top 20 upregulated circRNAs in five CRC and paired normal tissues. C Northern blotting and Sanger sequencing were used to characterize circGPRC5A. D–E CircGPRC5A was amplified from the CRC cell line cDNA (HCT8 and HT29), but not from gDNA, using divergent primers. F–G After RNase R treatment, qRT-PCR was used to confirm the expressions of circGPRC5A and GPRC5A mRNA in HCT8 and HT29 cells. H–I Using oligo dT and random 6-mers, we investigated the levels of circGPRC5A and GPRC5A mRNA expressions in HCT8 and HT29 cells. J–K The expressions of circGPRC5A and GPRC5A mRNA were assessed using qRT-PCR in HCT8 and HT29 cells following treatment with actinomycin D at the appropriate timepoint. The results of each assay, which were carried out three times, are displayed as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001
CircGPRC5A is upregulated in CRC and is correlated with clinicopathologic featuresThree CRC tissues and paired para-cancerous tissues were compared using the circGPRC5A FISH probe, which revealed that the expression of circGPRC5A in CRC tissue was higher than in matched adjacent normal tissues (Fig. 2A, Figure S1A–C). In addition, circGPRC5A was significantly upregulated in DLD-1, LOVO, SW620, HCT8 and HT29 cells compared with NCM460 cells (Fig. 2B). Extracted RNA from the 80 CRC tissues and corresponding adjacent normal tissues were subjected to qRT-PCR analysis to further confirm the presence of circGPRC5A in CRC tissues. CircGPRC5A was significantly upregulated in CRC tissues when compared to nearby normal tissues, which is consistent with our circRNA microarray data from CRC tissues (Fig. 2C).
Fig. 2
CircGPRC5A expression in CRC cell lines and correlations between circGPRC5A expression and CRC patients with clinicopathologic features. A FISH was used to investigate the expression of circGPRC5A in paired normal tissues and CRC tissues. B The qRT-PCR was used to confirm the expressions of circGPRC5A in NCM460 and CRC cell lines. C CircGPRC5A expressions in matched normal tissues and 80 CRC tissues. D-F T stage, lymph node status and Tumor size were considered when evaluating the expression of circGPRC5A. G The cellular localizations of circGPRC5A in HCT8 and HT29 cells were investigated using FISH. H-I The qRT-PCR was used to assess the expressions of circGPRC5A in nuclear and cytoplasmic RNA of HCT8 and HT29 cells. Based on data from three independent experiments, values are presented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001
Furthermore, high circGPRC5A tissue expression had a strong correlation with tumor size, lymph node status, T stage, and TNM stage (Fig. 2D–F) (Table S4). The distribution of circGPRC5A in HT29 and HCT8 cells was investigated using nuclear-cytoplasmic fractionation and FISH. In HCT8 and HT29 cells, the nuclear localization of circGPRC5A was slightly higher compared with cytoplasmic localization (Fig. 2G–I).
CircGPRC5A promotes CRC cell proliferation, migration and invasion in vitroTumor development and progression are significantly influenced by cell maintenance processes like proliferation and apoptosis resistance [25]. The expression of circGPRC5A was higher in HT29 and HCT8 cells compared with other CRC cell lines. Therefore, HT29 and HCT8 cells were used in subsequent studies. HT29 and HCT8 cells were transfected with si-circGPRC5A#1, si-circGPRC5A#2, si-circGPRC5A#3 and si-circGPRC5A#NC. qRT-PCR revealed that si-circGPRC5A#1 and si-circGPRC5A#2 gave a better knockdown efficiency compared with si-circGPRC5A#3 (Fig. 3A–C). We therefore used si-circGPRC5A#1 and si-circGPRC5A#2 to investigate the functional effects of circGPRC5A silencing in HT29 and HCT8 cells. CCK-8, colony formation, and EdU-incorporation revealed that silencing circGPRC5A reduced proliferation and colony formation (Fig. 3D–K).
Fig. 3
CircGPRC5A promotes CRC cell invasion, migration, and proliferation in vitro. A Schematic illustration of the siRNA targeted at the back-splice junctions of circGPRC5A. B–C After transfecting HCT8 and HT29 cells with siRNA or siNC, circGPRC5A expressions were assessed using qRT-PCR. D–H Colony formation and CCK-8 assays were used to measure proliferation after siRNA or siNC transfection into HCT8 and HT29 cells. I–K EdU assay (scalebar: 100 µm) was used to determine whether HCT8 and HT29 cells could proliferate after circGPRC5A silencing. L–N Transwell assays were performed to assess HCT8 and HT29 cell migration and invasion abilities after circGPRC5A silencing. O–Q Following circGPRC5A silencing, wound healing assays were carried out to measure migration and invasion of HCT8 and HT29 cells. Based on data from three independent experiments, values are presented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001
Previous results indicate that circGPRC5A expression is positively correlated with CRC patient lymph node status and TNM stage. Hence, to ascertain how circGPRC5A affects the migration and invasion of CRC cells, Transwell and wound healing assays were carried out. CircGPRC5A silencing significantly suppressed the migration and invasion abilities of HT29 and HCT8 cells (Fig. 3L–Q). We further explored the potential functional role of circGPRC5A by stably overexpressing circGPRC5A in CRC cell lines with lower circGPRC5A, HCT116 and SW480 cells, using lentiviral infection (Fig. 4A). Functionally, circGPRC5A overexpression significantly improved colony formation and cell proliferation as assessed by the CCK-8, colony formation, and EdU assays (Fig. 4B–G). The migration and invasion capacities of HCT116 and SW480 cells were also markedly enhanced with circGPRC5A overexpression (Fig. 4H–K). These data support that circGPRC5A can function as a tumor promotor to contribute to CRC development.
Fig. 4
CircGPRC5A promotes CRC cell invasion, migration, and proliferation in vitro. A CircGPRC5A overexpressions in HCT116 and SW480 were confirmed using qRT-PCR. B–G Following circGPRC5A overexpressions in SW480 and HCT116 cells, CCK-8, colony formation, and Edu assays were used to assess proliferation. H–K Transwell and wound healing assays were used to measure migration and invasion following circGPRC5A overexpressions in SW480 and HCT116 cells. Based on data from three independent experiments, values are presented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001. Following transfection of MS2-CP and circGPRC5A-MS2 into CRC cells, fluorescence and bright field images were obtained
CircGPRC5A interacts with PPP1CA and stabilizes PPP1CA proteinCircGPRC5A can promote bladder oncogenesis and metastasis by circGPRC5A-targeting peptide, and promote liver cancer progression by binding microRNAs [26, 27]. However, the mechanisms of circGPRC5A protein-binding are still unknown. Since circGPRC5A was distributed in both the cytoplasm and nucleus, we investigated protein-binding circRNA complexes influenced by circGPRC5A in CRC. First, we conducted proteomic profiling by using a circRNA pulldown assay with MS2-CP-Flag (Fig. 5A). CircGPRC5A-MS2 vector and MS2-CP-Flag were transfected into HCT116 cells, and qRT-PCR was used to verify the transfection efficiency (Fig. 5B,C). Then, a MS2-CP-Flag antibody was mixed with capture protein MS2-CP-Flag to further enrich for circRNA pulldown products. In the capture eluate, qRT-PCR revealed that circGPRC5A was highly abundant (Fig. 5D, E). Both assays successfully demonstrated the circRNA pulldown assay specificity. After mass spectrometry analysis of circRNA pulldown products, 33 candidate circGPRC5A-binding proteins were identified by comparing the circGPRC5A + MS2-CP-tansfected group with the circGPRC5A-MS2 + MS2-CP-transfected group. Mass spectrometry revealed that the value of protein matches and the exponentially modified protein abundance index were the highest for PPP1CA, which showed that the probability of circGPRC5A interacting with PPP1CA was the greatest. The function PPP1CA protein and its role in cancer progression, especially for CRC, is unknown.
Fig. 5
PPP1CA protein may be stabilized by circGPRC5A-PPP1CA binding. A RNA pulldown using the MS2-tagging system is outlined in a flow chart. B Following transfection of the expression plasmids circGPRC5A, NC, and circGPRC5A-MS2 into CRC cells, the relative expression of circGPRC5A was assessed using qRT-PCR. C Following transfections of MS2-CP and circGPRC5A-MS2 into CRC cells, fluorescence and bright field images were obtained. D The qRT-PCR was used to measure the enrichment of circGPRC5A in a complex with MS2-CP-Flag E MS2 combined protein in various groups in the RNA pulldown assay was examined using western blotting. F-G CircGPRC5A and PPP1CA were compared using the RIP assay to assess their relationship. The qRT-PCR was used to identify circGPRC5A expressions in the immunoprecipitates. H Immunofluorescence-FISH was used to examine co-localizations of circGPRC5A and PPP1CA in HT29 and HCT116 cells. I Following the overexpression or silencing of circGPRC5A, the relative expression of PPP1CA mRNA was assessed using qRT-PCR. J–L PPP1CA protein levels were assessed using western blotting and immunofluorescence after circGPRC5A was silenced or overexpressed. Values from three independent experiments are presented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001
Based on the above results, we decided to explore the relationship between circGPRC5A and PPP1CA. To further confirm the binding relationship between circGPRC5A and PPP1CA, a RIP assay was conducted to verify that circGPRC5A was specifically precipitated with PPP1CA in HT29 and HCT116 cells, in contrast to the control IgG group (Fig. 5F, G). Next, we used immunofluorescence-FISH assays to visualize and identify the colocalization of circGPRC5A and PPP1CA in both the cytoplasm and nucleus of HT29 and HCT116 cells (Fig. 5H). In addition, we explored how circGPRC5A affects PPP1CA by knocking down or overexpressing circGPRC5A in HT29 and HCT116 cells. We surprisingly found that PPP1CA mRNA levels remained unchanged with circGPRC5A expression manipulation (Fig. 5I). However, PPP1CA protein levels changed (Fig. 5J–L, Figure S6A). These results indicate that circGPRC5A can stabilize PPP1CA at the protein level.
In order to further explore the relationship between circGPRC5A and PPP1CA and verify that circGPRC5A can stabilize PPP1CA at the protein level, we conducted PPP1CA protein level detection in 40 pairs of cancer and adjacent samples, which revealed that PPP1CA expression was higher in cancer tissues than in adjacent tissues (Figure S2A, B). Using correlation analysis, we also found a positive correlation between PPP1CA and circGPRC5A, indicating that circGPRC5A affected the protein level of PPP1CA (Figure S2C).
CircGPRC5A inhibits PPP1CA and UBA1 binding and protects PPP1CA from proteasomal degradationUbiquitination and de-ubiquitination play important roles in the homeostasis of protein modification and biological activities, and are dynamic because of imbalances of ubiquitination and de-ubiquitination [28, 29]. The potential function of circGPRC5A to de-ubiquitinate PPP1CA protein, thereby stabilizing it, is unknown. To assess whether circGPRC5A affects PPP1CA protein at the post-translational level or protein synthesis level, the degradation rate of PPP1CA was measured using the protein synthesis inhibitor cycloheximide (CHX, 100 µg/ml). Interestingly, compared to the controls, CHX treatment resulted in lower PPP1CA protein with circGPRC5A silencing, and higher protein with circGPRC5A overexpression (Fig. 6A–D). This further demonstrates that circGPRC5A can affect PPP1CA protein at the post-translational level. Therefore, we used MG132, which is a proteasome inhibitor that can effectively inhibit the ubiquitination process of proteins via proteasomal degradation. For HT29 silencing of the circGPRC5A group, the expression of PPP1CA was partially reversed after using MG132. In contrast, for HCT116 overexpressing the circGPRC5A group, the expression of PPP1CA was stable after using MG132. The results revealed that circGPRC5A affected the expression of PPP1CA via proteasomal degradation (Fig. 6E and F, Figure S6B and C). A co-IP assay also demonstrated that silencing circGPRC5A in CRC cells increased PPP1CA protein poly-ubiquitination levels (Fig. 6G, Figure S6D). In contrast, overexpression of circGPRC5A significantly reduced PPP1CA protein poly-ubiquitination (Fig. 6H, Figure S6E). In conclusion, these results reveal that circGPRC5A can stabilize PPP1CA protein by preventing ubiquitin-mediated proteasomal degradation.
Fig. 6
CircGPRC5A stabilizes PPP1CA protein by inhibiting the binding between UBA1 and PPP1CA. A–D After silencing or overexpressing circGPRC5A in HT29 and HCT116 cells, respectively, transfected CRC cells were treated with CHX (100 µg/mL) at the indicated times, and PPP1CA protein was evaluated using western blotting. E–F After silencing or overexpressing circGPRC5A in HT29 and HCT116 cells, respectively, transfected CRC cells were treated with MG132 (20 µM) for 6 h and then PPP1CA was evaluated using western blotting. G–H An IP assay was used to detect the polyubiquitination level of PPP1CA after silencing or overexpressing circGPRC5A. The immunocomplexes were examined using western blotting with anti-Ub and anti-PPP1CA antibodies. I-J PPP1CA was immunoprecipitated with an anti-PPP1CA antibody, and IgG was used as the negative control. The immunocomplexes were analyzed using western blotting, and then the SDS-PAGE gel was stained with Coomassie Brilliant Blue. K Mass spectrometry analysis was used to analyze the products of immunocomplexes immunoprecipitated with anti-PPP1CA antibody. Ubiquitination related proteins are shown. L CircRNA inhibited the binding between UBA1 and PPP1CA. The immunocomplexes with an anti-PPP1CA antibody were examined by western blotting. M–N CircGPRC5A inhibited PPP1CA ubiquitin levels by inhibiting the binding between UBA1 and PPP1CA. Immunocomplexes with an anti-PPP1CA antibody were assessed using the indicated antibodies by western blotting
We did not find that related de-ubiquitination proteins interacted with circGPRC5A and PPP1CA in the RNA pulldown products. Hence, we speculate that circGPRC5A may interfere with binding between PPP1CA and related ubiquitination proteins. To test this hypothesis, an IP assay for PPP1CA was conducted and the products from the IP were analyzed using mass spectrometry. The presence of ten related ubiquitination proteins were confirmed (Fig. 6I–K). Among these ten related ubiquitination proteins, UBA1 and FBXO2 could directly bind to PPP1CA, while the remaining proteins typically affected the levels of protein ubiquitination by forming ubiquitination complexes [30, 31]. We hypothesized that circGPRC5A interfered with the binding of PPP1CA to UBA1 or FBXO2, thereby affecting PPP1CA protein levels. The ability of UBA1 and PPP1CA to bind changed after circGPRC5A was silenced or overexpressed, but not for FBXO2, according to an IP for PPP1CA (Fig. 6L, Figure S6F and G). Furthermore, the partial reversal of circGPRC5A causing the change of PPP1CA poly-ubiquitination level was observed through silencing or overexpressing UBA1 (Fig. 6M and N, Figure S6J–M). Collectively, circGPRC5A can interfere with the binding between UBA1 and PPP1CA and further stabilize PPP1CA protein levels.
PPP1CA and UBA1 are critical for circGPRC5A-mediated promotion of CRCPPP1CA is frequently overexpressed in tumors, and some studies have shown that correlations with clinicopathological traits are linked to the emergence of malignant tumors [17, 18]. CCK-8, colony formation, Transwell, and wound healing assays were conducted to investigated the effects of PPP1CA on circGPRC5A-mediated proliferation and migration of CRC cells. These experiments aimed to determine whether circGPRC5A stabilizes PPP1CA to influence the proliferation and migration of CRC cells. The partial reversal of circGPRC5 silencing-induced inhibitory effects on cell proliferation and migration was observed through the overexpression of PPP1CA in HT29 cells (Fig. 7A, B and E-G). In contrast, circGPRC5A overexpression-induced cell proliferation and migration in HCT116 cells could be partially mitigated with PPP1CA knockdown (Fig. 7C, D and H–J). These results support the notion that PPP1CA is critical for circGPRC5A-mediated CRC progression.
In previous studies, we reported that UBA1 was a key protein for circGPRC5A to stabilize PPP1CA. Therefore, we continued to explore the role of UBA1 on circGPRC5A-mediated promotion of CRC. By using CCK-8, colony formation, Transwell, and wound healing assays, partial reversal of circGPRC5 silencing-induced inhibitory effects on cell proliferation and migration were observed using UBA1 knockdown in HT29 cells (Figure S3A, B, E–G). In addition, circGPRC5A overexpression-induced cell proliferation and migration in HCT116 cells could be partially mitigated with overexpression of UBA1 (Figure S3C, D and H–J).
Fig. 7
PPP1CA is critical for circGPRC5A-mediated promotion of CRC. A–D CCK-8, colony formation, Transwell, and wound healing assays were conducted to investigate the effects of PPP1CA on circGPRC5A-mediated proliferation and migration of HT29 and HCT116 cells. E–J Experimental results of the effects of PPP1CA on circGPRC5A-mediated proliferation and migration of HT29 and HCT116 cells were quantified and presented using a bar chart. Values are shown as the mean ± SD based on three independent experiments. *P < 0.05; **P < 0.01; ***P < 0.001
Impact of circGPRC5A on YAP-Ser127 and YAP-Ser109 dephosphorylation through stabilization of PPP1CA proteinWe confirmed that circGPRC5A can stabilize PPP1CA, which is essential for circGPRC5A-mediated CRC progression. As a dephosphorylation protein, the imbalance of PPP1CA would inevitably lead to change of phosphorylation status, which could affect various biological behaviors. We hypothesized that circGPRC5A can lead to an imbalance in phosphorylation levels by stabilizing PPP1CA, thereby causing CRC progression (Fig. 8A). Phosphoproteomics was conducted to analyze the changes in phosphorylation states in circGPRC5A-silenced CRC cell lines. Phosphoproteomics revealed 4170 phosphopeptides, 4793 phosphorylation sites and 2390 phosphoproteins (Fig. 8B). Phosphoproteomics further demonstrated that PPP1CA mainly affected serine phosphorylation sites (89.61%), followed by threonine (9.7%) and lysine (0.69%) (Fig. 8C).
Fig. 8
CircGPRC5A can cause dephosphorylation of YAP-ser127 and YAP-ser109 by stabilizing the PPP1CA protein. A Western blotting was used to examine PPP1CA protein after silencing circGPRC5A. B Phosphoproteomics revealed 4,170 phosphopeptides, 4,793 phosphorylation sites, and 2,390 phosphoproteins. C The pie chart illustrates the types of phosphorylated amino acids from quantitative phosphoproteomics analysis. D Upregulated and downregulated protein phosphorylation levels from quantitative phosphoproteomics analysis. E Bubble chart depicting the deltaP_value and phosphorylation site number. F, G Western blotting was used to examine YAP phosphorylation after silencing and overexpressing PPP1CA or circGPRC5A. H–I The effects of YAP dephosphorylation via PPP1CA on circGPRC5A-induced changes were detected by western blotting
Phosphorylation enrichment results in the removal of most non-phosphorylated peptides, preventing accurate protein quantification. Therefore, based on the expression level and the number of phosphorylated peptides, a new variable called protein phosphorylation state value (deltaP_value) was defined. When deltaP_value ≥ 1, it indicates that the phosphorylated peptide in the protein is upregulated, and when deltaP_value ≤ − 1, it demonstrates that the phosphorylated peptide in the protein is downregulated. According to deltaP_value, we found upregulated phosphorylation levels of 383 proteins and downregulated phosphorylation levels of 201 proteins (Fig. 8D and E). Based on the deltaP_value and phosphorylation site number, we found that PPP1CA is most likely to lead to dephosphorylation of YAP and DBN1 proteins. YAP plays a pivotal role in cancer progression by changing phosphorylation [32]. DBN1 plays an important role in the formation of cell projections by changing expression [33, 34]. Although some studies have revealed that both YAP and DBN1 could participate in tumor progression, it is currently unclear whether changes in the phosphorylation status of DBN1 results in functional changes. Therefore, we inferred that circGPRC5A may lead to CRC progression by stabilizing PPP1CA and interfering with YAP phosphorylation levels.
The phosphoproteomics analysis revealed that YAP shows dephosphorylation at the following sites: Ser127, Ser138, Ser61, Ser109, Ser163/164 and Ser332. Dephosphorylation of YAP at Ser127 and Ser109 would cause YAP to enter the nucleus and bind to TEADs, thereby affecting cell proliferation [35, 36]. Phosphorylation of YAP-Ser61 may lead to transcriptional inhibition of YAP [37]. The role of other YAP phosphorylation sites is not clear. We next used co-IP assays to detect the total phosphorylation level of YAP. We found that silencing circGPRC5A or PPP1CA resulted in an increase in YAP phosphorylation levels. In addition, YAP phosphorylation of Ser127 and Ser109, but not Ser61, increased with silencing of circGPRC5A or PPP1CA (Fig. 8F, G, Figure S7A–J). We next investigated whether circGPRC5A can affect YAP phosphorylation levels by stabilizing PPP1CA. We found that the inhibiting effects on YAP dephosphorylation by silencing circGPRC5S was partially rescued after overexpression of PPP1CA in HT29 cells (Fig. 8H, Figure S7K–O). In HCT116 cells, circGPRC5A overexpression-induced YAP dephosphorylation could also be partially mitigated by knockdown of PPP1CA. (Fig. 8I, Figure S7P–T). We also extracted cytoplasmic and nuclear proteins separately and found that the change of YAP phosphorylation state led to changes in the distribution of YAP. PPP1CA-mediated dephosphorylation of YAP at Ser127 and Ser109 induced translocation of YAP in the nucleus (Figure S4A–D). These findings reveal that circGPRC5A may lead to CRC progression by interfering with YAP phosphorylation levels through stabilizing PPP1CA.
To further explore the important role of YAP on circGPRC5A/PPP1CA-mediated CRC progression, we demonstrated that the partial reversal of circGPRC5A/PPP1CA-mediated effects on cell proliferation and migration including CCK-8, colony formation, Transwell, and wound healing assays were observed using Verteporfin (YAP inhibitor, 10 ug/mL for 24 h) in HT29 and HCT116 cells (Figure S5A–J).
CircGPRC5A promotes the proliferation and migration of CRC cells in vivoTo verify the effect of circGPRC5A on CRC cell proliferation in vivo, circGPRC5A-knockdown and circGPRC5A-overexpression HT29 and HCT116 cells were xenografted subcutaneously into nude mice. The size of the xenograft tumors was measured on days 5, 10, 15, 20, and 25 after tumor cell injection. The circGPRC5A-overexpressing HCT116 tumors were the largest by volume and weight with the fastest tumor growth compared with the vector control group. On the other hand, knocking down circGPRC5A in HT29 cells resulted in smaller tumors by volume and weight, and the tumors were slower-growing (Fig. 9A–D).
Fig. 9
CircGPRC5A induces the growth of CRC cells in vivo. A and B Images of subcutaneous xenograft tumors. C After xenograft implantation, the xenograft tumor growth curve was assessed every 5 days. D Subcutaneous xenograft tumor weights. E The effect of knocking-down circGPRC5A on intrahepatic metastasis. F Images of hematoxylin-and-eosin-stained intrahepatic metastases. G Ki-67 and PPP1CA staining and IHC analysis of subcutaneous xenograft tumors. H Diagram illustrating the role of circGPRC5A in CRC
Next, we explored the effect of circGPRC5A on CRC cell metastases. Female nude mice were injected with stably-transfected HCT8 cells with either circGPRC5A knockdown vector or the empty vector control. In contrast to the control group, we discovered that the circGPRC5A knockdown group had fewer metastatic nodules (Fig. 9E, F). IHC staining for Ki-67 and PPP1CA was performed on subcutaneous xenograft tumor tissues. Compared with the control group, Ki-67 and PPP1CA expression increased in tumors from the circGPRC5A-overexpressing group and decreased in tumors from the circGPRC5A-knockdown group (Fig. 9G, Figure S8A–C). Consistent with the in vitro experiments, xenograft tumor experiments revealed that circGPRC5A could promote CRC growth.
Comments (0)