
Remember me




Considering the limited research on the chemical composition of S. melanocentra, the concentrated methanolic extract was initially analyzed using an analytical column, ReproSil-Pur C18 AQ. The chromatogram depicting the analysis is presented in Fig. 1A. As observed in Fig. 1A, an index component was detected in the extract. However, the ReproSil-Pur C18 AQ analytical column did not provide optimal separation for the index component, resulting in a peak with poor symmetry and a nonideal parcel-like shape (highlighted by the red box in Fig. 1A). This indicates that the ReproSil-Pur C18 AQ column may not be the most suitable choice for achieving high-purity separation of the index component in S. melanocentra. Regarding the effectiveness of polyamide chromatography in separating flavonoids, phenolics, quinones, and polar compounds, such as alkaloids and steroids, from nonpolar compounds [31, 32], it was deemed a more suitable option. Furthermore, the large separation volume of the polyamide column makes it appropriate for preparative separation. Subsequently, the mixed sample was crushed, sieved, and weighed, resulting in a total sample weight of 120.5 g. Based on calculations, the yield of the total sample was determined to be 10.3%, equivalent to 20.5 g.
Fig. 1
Analytical chromatogram on a Reprosil-Pur C18 AQ analytical column of the S. melanocentra methanol extract (A) and separation chromatogram of the S. melanocentra polyamide mixture (B)
Polyamides are macromolecular substances that result from the polymerization of amides and contain multiple amide groups in their molecular structure [33]. In this study, a cost-effective MPLC technique utilizing polyamide was employed to isolate bergenin from the methanol extract of S. melanocentra. During the experimental process, a preparative liquid chromatograph equipped with a high-performance medium-pressure column (49 × 460 mm) filled with polyamide was employed. Additionally, a small medium-pressure column (49 × 100 mm) was used to facilitate the loading of a mixture comprising dried polyamide and the sample (60.0 g) for the preparation of dry-loaded samples. The resulting chromatogram of the preparation is depicted in Fig. 1B. Following two enrichment cycles using an acetonitrile-water eluent, five fractions (Fr1, Fr2, Fr3, Fr4, and Fr5) were collected. The results demonstrated the efficacy of polyamide MPLC in achieving the efficient preparation of active ingredients, thus highlighting it as a favorable method for sample separation.
After two repeated separations, the collected target fraction was combined and subjected to concentration, resulting in the obtainment of 1.36 g of the desired sample (Fr2) with a recovery rate of 6.8%. The HPLC chromatograms of the methanol extract of S. melanocentra and the Fr2 sample were then analyzed and compared using the ReproSil-Pur C18 AQ analytical column. The corresponding results are presented in Fig. 2A and B. The analysis showed that the primary compound present in the crude extract was significantly enriched in the Fr2 fraction. Notably, Fr2 exhibited a single peak with excellent symmetry when subjected to analysis on the ReproSil-Pur C18 AQ analytical column, indicating a sufficiently high level of purity. These findings suggest that polyamide and C18 columns possess complementary selectivity and can be effectively combined for the separation of bergenin.
Fig. 2
HPLC analysis of the S. melanocentra methanol extract (A), Fr2 (B) on the Reprosil-Pur C18 AQ analytical column, purity analysis comparison figures of the isolated main fraction Fr2 on the ReproSil-Pur C18 AQ, XCharge C18, XAmide, and Click XIon analytical column (C) and chemical structures of the isolated compound (D)
To further validate the purity of Fr2, additional analysis was conducted using an RP column (XCharge C18) and HILIC columns (XAmide and Click XION). The corresponding analytical chromatogram is presented in Fig. 2C. Upon examination of these chromatographic columns, it was observed that the main component Fr2 exhibited a purity level exceeding 99%. Retention mechanisms in reversed-phase liquid chromatography (RPLC) and hydrophilic interaction liquid chromatography (HILIC) differ due to varying principles [34, 35]. The ReproSil-Pur C18 AQ analytical column is bonded with ultrapure silica gel and specifically designed for hydrophilic and polar compounds, offering improved retention and selectivity. It demonstrates excellent pH stability and batch repeatability. The XCharge C18 analytical column employs electrostatic control technology, allowing for the regulation of hydrophobicity and electrostatic properties on the material surface by balancing positive and negative charges. This enables effective separation of analytes. The XAmide column, featuring a neutral amide-bonded phase, exhibits remarkable hydrophilicity, thereby overcoming issues related to the acidity and inhomogeneity of silanol groups found on pure silica gel surfaces. It provides an alternative to commonly used amino and silica gel columns by avoiding potential electrostatic effects. The XAmide column facilitates good peak shapes and separations for various strongly polar compounds [36]. Moreover, the Click XIon analytical column possesses surface hydrophilicity and controllable surface electrostatic effects, allowing for the separation of highly polar and charged hydrophilic compounds [37, 38]. Considering the complementary selectivity of the RP columns (ReproSil-Pur C18 AQ and XCharge C18) and HILIC columns (XAmide and Click XIon), the purity of the main component Fr2 from S. melanocentra was thoroughly confirmed.
The obtained ESI-MS, 1 H NMR, and 13 C NMR spectra were compared with published literature to determine the structure of the main compound, Fr2 (Fig. 2D). The structural identification of Fr2 is presented in Fig. 3A-D of the supplementary information. Based on the spectral data, the target compound was identified as bergenin. Compound Fr2 (bergenin) was obtained as a white powder weighing 1.36 g. The ESI-MS spectrum exhibited peaks at m/z 351.19 ([M + Na]+) and m/z 327.16 ([M − H]−), while the calculated mass for C14H16O9 was m/z 328.08. The 1 H NMR spectrum (600 MHz, MeOH-d4) displayed signals at 7.08 (1 H, s, H-7), 4.95 (1 H, d, J = 10.5 Hz, H-10b), 4.04 (2 H, m, H-4a, H-4), 3.90 (3 H, s, H-12), 3.80 (1 H, t, J = 7.6 Hz, H-2), 3.68 (2 H, m, H-11), and 3.42 (1 H, m, H-3). The 13 C NMR spectrum (151 MHz, MeOH-d4) exhibited peaks at 165.8 (C-6), 152.4 (C-8), 149.5 (C-10), 142.3 (C-9), 119.5 (C-6a), 117.3 (C-10a), 111.0 (C-7), 83.1 (C-2), 81.5 (C-4a), 75.6 (C-4), 74.3 (C-10b), 71.9 (C-3), 62.7 (C-11), and 60.9 (C-12). The obtained ESI-MS, 1 H NMR, and 13 C NMR data were consistent with the literature data for bergenin [39].
Fig. 3
ESI-MS and NMR of the isolated compound Fr2 (bergenin). \(+\)ESI mass of compound Fr2 (A), \(-\)ESI mass of compound Fr2 (B), 1 H NMR spectrum (600 MHz) of compound Fr2 (C), and 13 C NMR spectrum (151 MHz) of compound Fr2 (D)
Pharmacological effects of bergenin against hyperlipidemia based on the zebrafish modelAcute toxicity test results of bergeninThe outcomes of the acute toxicity study revealed that petroselin at various concentrations (0, 10, 33.3, and 100 μM) did not induce toxicity in zebrafish. The mortality rate of zebrafish larvae was assessed at different bergenin concentrations (0, 10, 33.3, and 100 μM), as depicted in Fig. 4A and B. Zebrafish were deemed deceased if no visible activity was observed in the petri dish during the experiment and touching the tail did not elicit a response. There was no notable variation in the mortality rate relative to the exposure concentration of zebrafish after 96 h of exposure to petroselin in the experiment. Moreover, the survival and hatching rates of zebrafish larvae in the 0, 10, 33.3, and 100 μM bergenin treatment groups consistently exceeded 80%. Based on these results, doses of 5 μM and 15 μM were chosen for subsequent experiments, ensuring that they remained below the concentration of 33.3 μM.
Fig. 4
Effect of different concentrations of bergenin on embryonic mortality and hatching rate of zebrafish. Figure A shows the survival rate, and Figure B shows the hatching rate
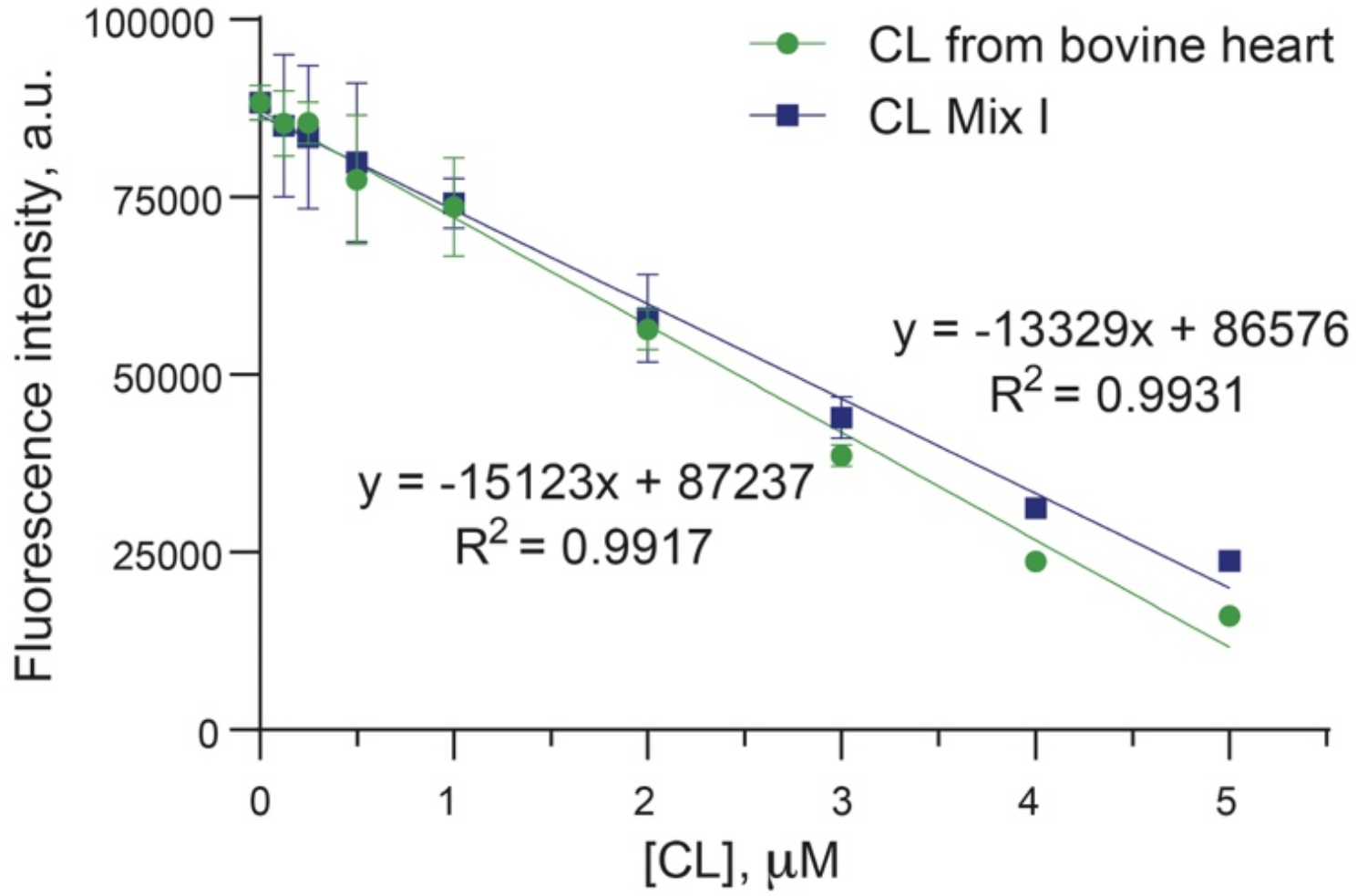
Effect of bergenin on lipid levels in zebrafishIn our investigation, we explored the hypolipidemic impact of bergenin on a zebrafish model of hyperlipidemia induced by a high-fat diet. Figure 5 A-D depict the lipid profile of zebrafish larvae. In the model group, there were significantly heightened levels of TG (P < 0.05), TC (P < 0.01), and LDL-c (P < 0.01), accompanied by significantly reduced levels of HDL-c compared to the blank control group (P < 0.05). However, in the bergenin-treated group, there were significant reductions in the levels of TG, TC, and LDL-c, coupled with a significant increase in HDL-c. The changes in the levels of TG, TC, LDL-c, and HDL-c exhibited a dose-dependent relationship when compared with the model control group. Specifically, as the concentration of bergenin increased to 15 μM, the levels of TG, TC, and LDL-c in zebrafish decreased by 12.49% (P < 0.01), 4.27% (P < 0.01), and 2.96% (P < 0.01), respectively. Furthermore, the HDL-c levels in zebrafish increased by 0.29% (P < 0.01) when the concentration of bergenin was elevated to 15 μM. Notably, the higher concentration of bergenin exhibited a superior therapeutic effect compared to fenofibrate in reducing TC and LDL-c levels. Hyperlipidemia is a common disorder characterized by elevated plasma TG, TC, and LDL-c levels as well as decreased HDL-c levels. LDL-c plays a vital role in transporting TC to peripheral tissues, but excessive LDL-c and TC can lead to their deposition on arterial walls, resulting in endothelial cell damage, increased vascular permeability, and the development of atherosclerosis. This process involves the entry of plasma lipoproteins into the arterial intima, the recruitment and differentiation of monocytes into macrophages, and the formation of foam cells, which ultimately contribute to the formation of fibrous and atheromatous plaques, leading to cardiovascular diseases such as atherosclerosis [40, 41]. On the other hand, HDL-c facilitates the transport of excess cholesterol from peripheral tissues to the liver for metabolism and exhibits potent anti-inflammatory effects, thereby playing a protective role against atherosclerosis [42]. Lowering TG, TC, and LDL-c levels while increasing HDL-c levels is crucial for the prevention and treatment of cardiovascular diseases. The findings indicate that bergenin possesses effective regulatory properties on lipid metabolism disorders by reducing TG, TC, and LDL-c levels and increasing HDL-c levels. These results highlight the potential of bergenin in managing lipid disorders and preventing cardiovascular diseases.
Fig. 5
Effect of bergenin on blood lipid levels in high-fat zebrafish. Panels A, B, C, and D represent the contents of TG, TC, LDL-c, and HDL-c, respectively. Ffb: fenofibrate; Ber: bergenin. Values are presented as the mean ± SD. *P < 0.05, **P < 0.01 vs. the control group; #P < 0.05, ##P < 0.01 vs. the model group
Effect of bergenin on overall lipid accumulation in zebrafishThe solubility of Oil Red O dye is higher in tissues and cells compared to its original solvent, enabling its binding to TG and the formation of lipid droplets with an orange‒red appearance. Juvenile zebrafish, characterized by relatively transparent bodies with minimal melanin accumulation, allow for clear observation of lipid accumulation status under a microscope after staining. The staining results are depicted in Fig. 6A-E. In the control group, minimal Oil Red O staining was observed in the liver and tail vessels of zebrafish juveniles. In contrast, the high lipid model group displayed prominent orange‒red staining in these areas, indicating successful establishment of the zebrafish hyperlipidemic model with the accumulation of large lipid droplets. Lipid droplets were significantly reduced in the positive control (fenofibrate) and bergenin-treated groups compared to the model group. Remarkably, orange‒red lipid droplets in the liver and caudal vessels of zebrafish larvae decreased significantly and appeared lighter in color with increasing doses of petroselin. Notably, the high concentration of bergenin (15 μM) demonstrated a more pronounced lipid-lowering effect than fenofibrate. The effect of 15 μM petroselin in reducing lipid deposition in zebrafish was comparable to that of lipid staining in zebrafish in the blank control group. These findings provide additional evidence that bergenin effectively mitigates lipid deposition in the liver and caudal vessels of high-fat zebrafish larvae, highlighting its potential as a lipid-lowering agent.
Fig. 6
The effect of bergenin on overall lipid accumulation in high-fat zebrafish. (A): HE staining results in the control group; (B): HE staining results in the model group; (C) HE staining results in the 1.5 μM fenofibrate group; (D) and (E): HE staining results in the 5 and 15 μM bergenin groups. The upper figure represents a magnification of 20×, the middle figure represents a magnification of 40×, and the lower figure is a partial magnification of the zebrafish liver area (red box). Ffb: fenofibrate; Ber: bergenin. Values are presented as the mean ± SD. *P < 0.05, **P < 0.01 vs. the control group; #P < 0.05, ##P < 0.01 vs. the model group
Effects of bergenin on the behavior of high-fat zebrafishExcessive lipid accumulation in the body adversely affects physical activity. The consumption of a high-fat diet disrupts the secretion of muscle factors and interferes with glucolipid metabolism, leading to obesity and impairing overall body movement. The utility of behavioral responses as indicators of motor neuron function in organisms has been established [43]. Moreover, studies have investigated the impact of lipid-lowering compounds on motor behavior in the zebrafish hyperlipidemic model system [44, 45]. To evaluate the effects of different treatments, behavioral tests were conducted on zebrafish in various experimental groups, and the findings are illustrated in Fig. 7A-F. Zebrafish in the high-fat model group exhibited a significantly reduced total swimming distance compared to the blank control group, indicating compromised activity levels. Compared to the model group, zebrafish treated with bergenin (5 μM) and fenofibrate (1.5 μM) showed increased swimming distances, although the differences were not statistically significant. However, zebrafish in the 15 μM bergenin group demonstrated a significant increase in total swimming distance (P < 0.05). These results suggest that bergenin has the potential to enhance the impaired behavior of zebrafish caused by a high-fat diet. Furthermore, the concentration of bergenin showed a positive correlation with the observed improvement in behavioral performance.
Fig. 7
Effect of bergenin on the behavior of high-fat zebrafish. Figures A, B, C, D, and E represent the behavioral trajectory of each group of zebrafish, and F represents the total distance traveled by zebrafish in 15 s. Ffb: fenofibrate; Ber: bergenin. Values are presented as the mean ± SD. *P < 0.05, **P < 0.01 vs. the control group; #P < 0.05, ##P < 0.01 vs. the model group
Mechanism of action of bergenin against hyperlipidemiaCholesterol, a vital lipid in living organisms, is predominantly taken up by macrophages, serving as a major pathway for its uptake in the body. Macrophage lipid metabolism disorders encompass irregular cholesterol uptake and conversion, aberrant cholesterol reversal and extracellular transport, and abnormal expression of genes involved in lipid metabolism regulation [46, 47]. To explore the underlying mechanism of the effect of bergenin on improving hyperlipidemia, the present study employed qRT‒PCR to assess the expression levels of various genes, including FASN, SREBF1, LPL, HMGCRα, RORα, LDLRα, IL-4, IL-1β, and TNF, across different experimental groups. FASN, a complex enzyme system consisting of seven enzymes encoded by the same gene, predominantly resides in liver tissue. Its activity directly influences the liver’s capacity for synthesizing fatty acids, playing a critical role in regulating fat accumulation in animals [42]. Acting as an upstream transcription factor, SREBF1 governs the expression of FASN and exerts an impact on lipid aggregation by regulating fatty acid synthesis, thereby promoting lipogenesis and maintaining lipid homeostasis [42]. Lipoprotein lipase (LPL) serves as a pivotal rate-limiting enzyme in lipid catabolism, facilitating the breakdown of triglycerides into free fatty acids and mono-fatty acid triglycerides to provide the body with energy through oxidative breakdown [48]. 3-Hydroxy-3-methylglutaryl coenzyme A reductase (HMGCR), a vital component of human lipid synthesis, plays a crucial role in regulating cholesterol synthesis and metabolism. As cholesterol synthesized in the liver serves as the primary source of cholesterol in the body, HMGCR acts as the rate-limiting enzyme catalyzing the conversion of 3-hydroxy-3-methylglutaryl coenzyme A to mevalonate during cholesterol synthesis. Inhibiting HMGCR activity in the liver can reduce cholesterol synthesis and help regulate lipid metabolism disorders [49]. Low-density lipoprotein receptor alpha (LDLRα) is a hepatic receptor responsible for mediating the endocytosis of cholesterol-rich LDL particles, thereby maintaining optimal LDL plasma levels. Retinoic acid receptor-related orphan receptor alpha (RORα) plays a pivotal role in maintaining lipid metabolism homeostasis by regulating the transcriptional activity of target genes involved in triacylglycerol metabolism. Additionally, RORα serves as a negative regulator of inflammation [50]. Interleukin-4 (IL-4) is a growth factor secreted by T cells that functions as an anti-inflammatory factor with immunosuppressive effects on the body [51]. Interleukin-1 beta (IL-1β) and tumor necrosis factor alpha (TNF-α) are essential cytokines in the body and are primarily produced by active monocytes and vascular endothelial cells. They exert diverse biological effects and play a role in various pathological changes associated with inflammatory lesions in the body [52, 53].
The results of the experiment are presented in Fig. 8A-I. In comparison to the blank control group, the high-fat model group exhibited significantly increased expression levels of FASN, HMGCRα, SREBF1, LDLR, RORα, IL-1β, and TNF mRNA. Conversely, the expression levels of LPL and IL-4 mRNA were significantly downregulated. In the bergenin treatment group, compared to the model group, the expression levels of SREBF1, HMGCRα, and IL-1β mRNA were reduced, with statistically significant differences observed under 15 μM bergenin administration. The expression levels of FASN, RORα, LDLRα, and TNF mRNA were significantly downregulated under 5 μM and 15 μM bergenin administration, showing statistically significant differences that were concentration dependent. After bergenin treatment, the expression level of IL-4 mRNA was significantly upregulated. However, bergenin did not affect the expression level of LPL mRNA. These experimental findings indicate that bergenin can modulate lipid synthesis, cholesterol metabolism, and the abnormal expression of inflammation-related genes. The results suggest that bergenin primarily reduces lipid deposition by inhibiting lipid synthesis rather than promoting lipolysis via the SREBF1 and FASN-related signaling pathways. Additionally, bergenin ameliorates abnormal lipid metabolism by inhibiting RORα expression, thereby regulating HMGCRα, LDLRα, IL-4, IL-1β, and TNF expression to inhibit cholesterol synthesis and mitigate the body’s inflammatory response.
Fig. 8
Bergenin impacts the gene expression of FASN, SREBF1, HMGCRα, LPL, LDLRα, RORα, IL-4, IL-1β, and TNF mRNA in high-fat diet-fed zebrafish. FASN mRNA expression (A), SREBF1 mRNA expression (B), HMGCRα mRNA expression (C), LPL mRNA expression (D), LDLRα mRNA expression (E), RORα mRNA expression (F), IL-4 mRNA expression (G), IL-1β mRNA expression (H), and TNF mRNA expression (I). Ffb: fenofibrate; Ber: bergenin. Values are presented as the mean ± SD. *P < 0.05, **P < 0.01 vs. the control group; #P < 0.05, ##P < 0.01 vs. the model group
Molecular dockingMolecular docking is a technique used to identify the active site where a small molecule ligand interacts with a target protein molecule, allowing them to form a stable and energetically favorable binding conformation. The binding energy value reflects the strength of the interaction between the compound and the protein receptor, with lower binding energy values indicating higher binding activity [54]. To corroborate the outcomes of the RT‒PCR experiments, molecular docking was employed to visualize the binding interactions of bergenin with its key protein targets, as illustrated in Fig. 9A-I. Bergenin docks into a position analogous to the original ligand, occupying the active pocket formed by the amino acid residues of the protein. The binding energies of bergenin to the eight target receptors (FASN, SREBF1, HMGCRα, RORα, LDLRα, IL-4, IL-1β, and TNF) were all ≤ -6.0 kcal/mol, as indicated in Table 2. Specifically, the binding energies of bergenin (Fr2) to IL-4, LDLRα, and RORα were − 7.53, -7.48, and − 7.10 kcal/mol, respectively, indicating their strong binding activity, as the values were lower than − 7.0 kcal/mol. Expanding on this, the potent binding interactions between bergenin and IL-4 are primarily facilitated through robust hydrogen bonding interactions with LEU-7 and ASN-15 amino acid residues, along with the formation of Pi-alkyl, electrostatic, and π-π interactions with LEU-7, LEU-14, ILE-11, TYR-124, and LYS-12, residues. The interactions between bergenin and LDLRα exhibit Pi-Sigma interactions with PRO-639 residues, in addition to hydrogen bonding (PRO-639, HIS-591, SER-564, ARG-495, GLU-498, ALA-637) and Pi-alkyl (VAL-644, ALA-637) forces with the residues. Similarly, hydrogen bonding (THR-485, GLY-482, ARG-478, ALA-358) and Pi-alkyl (ARG-478, CYS-481, LEU-361) forces are formed between bergenin and RORα with the residues. Conversely, bergenin demonstrated a binding energy of only − 4.06 kcal/mol with the LPL protein. A docking binding energy below − 5.0 kcal/mol suggests that the compound exhibits some binding activity with the target protein, while a docking binding energy below − 7 kcal/mol indicates robust binding activity between the compound and the target protein.
Fig. 9
Molecular docking visual analysis of bergenin binding to FASN, SREBF1, HMGCRα, LPL, LDLRα, RORα, IL-4, IL-1β, and TNF. (A), (B), (C), (D), (E), (F), (G), (H), and (I) correspond to the binding models of bergenin with FASN, SREBF1, HMGCRα, LPL, LDLRα, RORα, IL-4, IL-1β, and TNF, respectively
Table 2 Intermolecular interactions between bergenin and FASN, SREBF1, HMGCRα, LPL, LDLRα, RORα, IL-4, IL-1β, and TNFTherefore, the molecular docking results of bergenin with these target proteins are consistent with the findings from the qRT‒PCR experiments conducted in the zebrafish model. In the in vivo experiments investigating the effects of bergenin on high-fat zebrafish, the qRT‒PCR results revealed that a low concentration (5 μM) of bergenin effectively regulated the mRNA expression of IL-4, LDLRα, and RORα. Furthermore, a high concentration (15 μM) of bergenin effectively regulated the expression of FASN, SREBF1, HMGCRα, IL-1β, and TNF mRNA. However, neither low nor high concentrations of bergenin showed a significant effect on the expression of LPL mRNA. Molecular docking serves to explore the theoretical level of specific action modes and binding conformations between small drug molecules and macromolecular targets, while qRT‒PCR experiments in the zebrafish model investigate the actual gene level. The mutual validation of these two approaches provides a scientific basis for understanding the mechanism of action of bergenin against hyperlipidemia. In conclusion, this study showed that bergenin exerts an effect on lipid metabolism in zebrafish fed a high-fat diet, and its mechanism of action may involve SREBF1-, FASN-, and RORα-related signaling pathways. However, further investigation is needed to elucidate the specific molecular mechanisms underlying the action of bergenin.
Comments (0)