
記住我
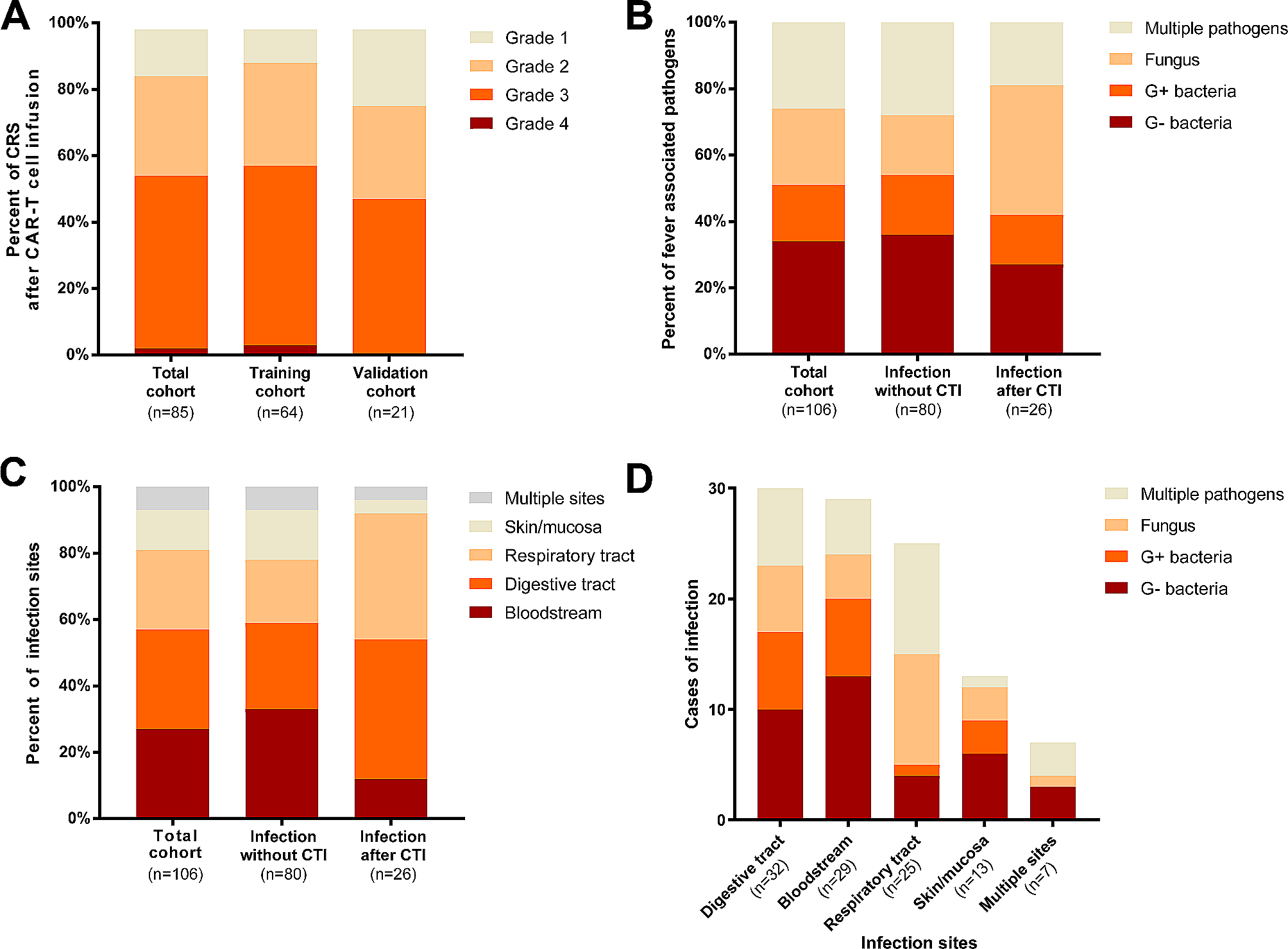






To induce acquired resistance to DOT1L inhibition, the KMT2A::AFF1+ ALL cell line SEM [22] was exposed to increasing concentrations of the first-in-class DOT1L inhibitor pinometostat for 14 weeks (Fig. 1A). Next, cells were cultured in the absence of drug before evaluating potential changes in pinometostat-induced cytotoxicity. Exposure of maternal SEM cells to 50 µM pinometostat for 1 week reduced the percentage of viable cells to ~ 35%.
Fig. 1
Establishment of acquired resistance to DOT1L inhibition in KMT2A-rearranged ALL cells. A. Graphic overview of acquired resistance induction to DOT1L inhibition in KMT2A::AFF1 + B-cell ALL (SEM) leading to pinometostat-resistant cells (SEMPINO_RES). B. Viable cell percentage of SEM and SEMPINO_RES cells in the absence (-) or presence (+) of 50µM pinometostat for 7 days, normalized to cells cultured without pinometostat. Data present the mean +/- the standard deviation (SD) derived from 2 biological replicates. C. IC50 values of viable cells of the indicated cell line models determined using six drug concentrations (0-100 µM) for 14 days. The data illustrates the mean +/- SD from 4 biological replicates, each comprising 3 technical replicates. D. Immunoblot images of H3K79me2 and total histone H3 in SEM and SEMPINO_RES cells cultured with or without 50 µM pinometostat for 7 days. E. Quantification of H3K79me2 protein expression using densitometry analysis normalized against total histone H3 expression. Data represent fold-changes normalized against untreated SEM cells for 2 biological replicates. F. mRNA expression of DOT1L, KMT2A:AFF1, and HOXA9 determined by qRT-PCR analysis, and G. viable cell percentage in SEM and SEMPINO_RES at day 2 and day 4 after siRNA-mediated knockdown (KD) relative to non-silencing controls (NSCs). Data of 2 biological replicates ± SD, *p < 0.05, **p < 0.005, ***p < 0.0005, ****p < 0.0001. (H) Immunoblot images of DOT1L, H3K79me2, and GAPDH protein expression in SEM and SEMPINO_RES cells at day 4 following siRNA-mediated KD of DOT1L, and corresponding quantification of (I) DOT1L or (J) H3K79me2 protein expression relative to GAPDH using densitometry analysis.
Differences were statistically evaluated using unpaired t-tests
In contrast the viability of SEM cells that underwent prolonged exposure to increasing pinometostat concentrations was hardly affected (Fig. 1B). This pinometostat-resistant daughter line, designated as SEMPINO_RES, revealed a 34-fold higher 14-day-IC50 value as compared to maternal SEM cells (Fig. 1C), indicating that SEMPINO_RES became highly resistant. An additional model of intermediate resistance was established in SEM cells (i.e., SEMPINO_INT) by prolonged exposure to 4.5 µM pinometostat for 7 weeks, leading to a 16-fold higher IC50 (Fig. 1C).
Similar to SEMPINO_RES, we also induced pinometostat resistance in the KMT2A::AFF1+ ALL cell line RS4;11. With a mean 14-day-IC50 value of ~ 10 µM, representing pinometostat concentrations well above maximum achievable plasma levels in pinometostat-treated patients, [15, 16, 23] maternal RS4;11 is more resistant than SEM (Fig. 1C). Two emerging pinometostat-resistant RS4;11 daughter lines, i.e., RS4;11PINO_RES#1 and RS4;11PINO_RES#2, showed IC50 values of 33–45 µM (Fig. 1C).
To assess the inhibitory effects of pinometostat on DOT1L-mediated methyltransferase activity, the levels of H3K79 di-methylation (H3K79me2) were determined by immunoblot analysis. Interestingly, the levels of H3K79me2 in SEM and SEMPINO_RES were comparable and pinometostat was able to substantially reduce the levels of H3K79me2 equally in both cell lines (Fig. 1D-E). Hence, despite persistent inhibition of DOT1L-mediated H3K79 methylation, cell viability in SEMPINO_RES is no longer affected, suggesting that these cells became largely independent of H3K79 methylation induced by DOT1L.
Next, we assessed whether changes in global histone modifications had occurred between SEM and SEMPINO_RES. For this we used Mod Spec®, a mass spectrometry-based measurement for the relative abundance of over 80 distinct histone marks. This analysis confirmed no differences in the levels of H3K79 mono-, di-, and tri-methylation (i.e., H3K79me1, H3K79me2, and H3K79me3, respectively) between SEM and SEMPINO_RES, and showed equal reduction of these histone marks upon pinometostat exposure (Figure S1). Moreover, these data demonstrated that the global landscape of histone modifications between SEM cells and SEMPINO_RES largely remained similar. The only histone modification that is downregulated in response to pinometostat exposure appeared to be H3K79 methylation, demonstrating the specificity of this agent.
In KMT2A-rearranged acute leukemias, the KMT2A fusion protein is considered to be the main oncogenic driver and loss of DOT1L was shown to specifically decrease KMT2A fusion-driven transcriptional programs, including the expression of HOXA9. [7] Therefore, we asked whether acquired resistance to DOT1L inhibition was accompanied by an altered dependency on DOT1L, KMT2A::AFF1 and/or HOXA9. Therefore SEM and SEMPINO_RES cells were subjected to siRNA-mediated knockdown of these genes, resulting in significant reductions in mRNA expression of ~ 75–80% for DOT1L and KMT2A::AFF1, and ~ 50–65% for HOXA9, relative to non-silencing controls (NSCs) (Fig. 1F). Validation on the protein level confirmed a reduction of DOT1L of ~ 90% in SEM and of > 90% in SEMPINO_RES (Fig. 1H,I). Knockdown of DOT1L was accompanied by a reduction of H3K79me2 of ~ 90% in SEM, and of ~ 40% SEMPINO_RES (Fig. 1H, J). Surprisingly, knockdown of DOT1L and KMT2A::AFF1 resulted in similar reductions in cell viability in both SEM and SEMPINO_RES. For HOXA9 suppression the effects on cell viability in SEMPINO_RES appeared to be somewhat delayed (Fig. 1G). This suggests that although SEMPINO_RES cells became less sensitive to inhibition of H3K79 methylation in terms of leukemic cell survival, these cells remained dependent on the physical presence of proteins known to be important in KMT2A-mediated leukemogenesis, including DOT1L.
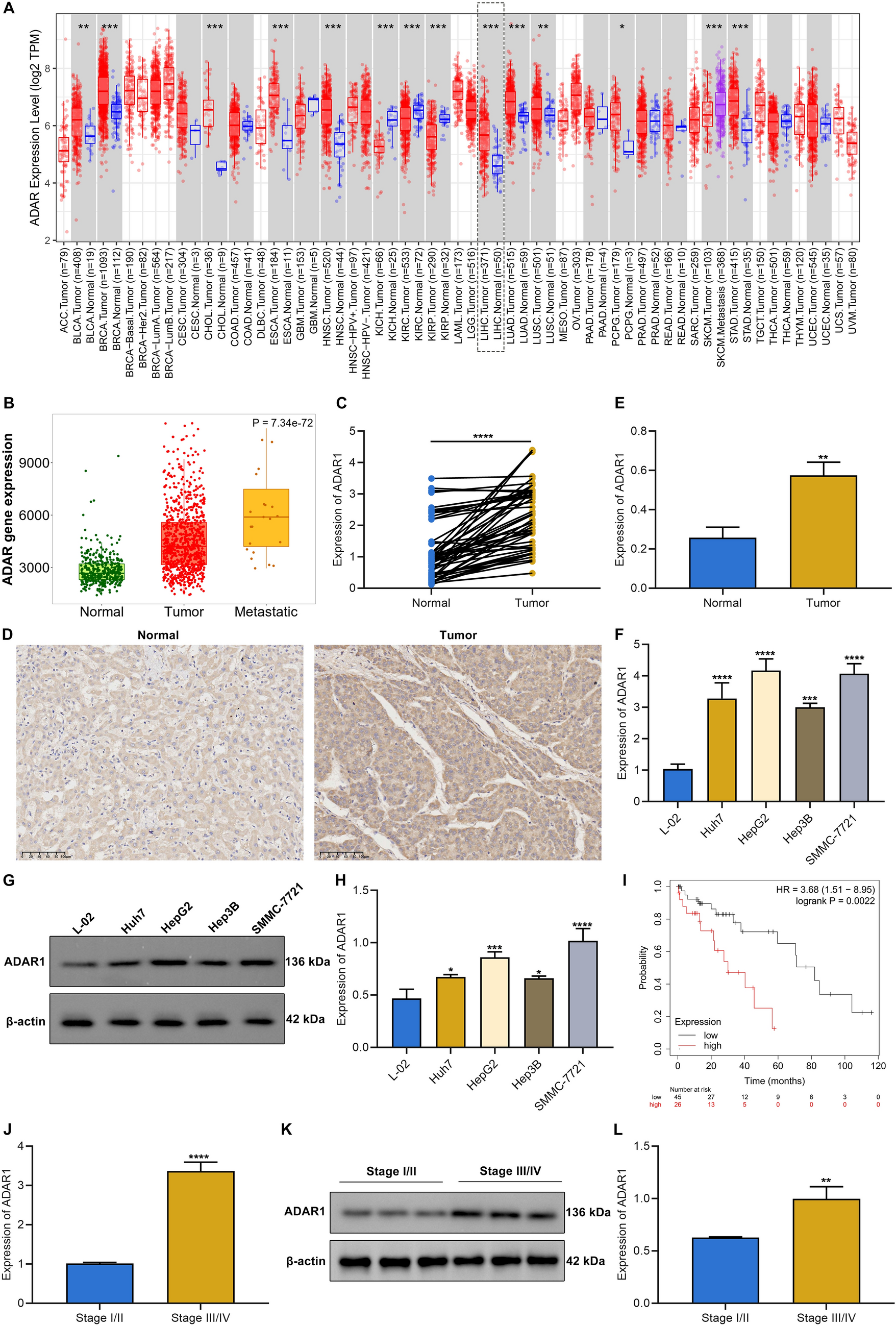
Acquired resistance to DOT1L inhibition leads to selective loss of KMT2A-fusion driven gene expressionNext, we performed RNA- and ChIP-seq for KMT2A, AFF1, H3K4me3, H3K79me2, and H3K27ac, as well as ATAC-seq on SEM and SEMPINO_RES cells cultured in both the absence and presence of 50 µM pinometostat for 7 days. Interestingly, as assessed by ChIP-seq, there are very few observable differences in the global profiles of KMT2A, AFF1 or H3K79me2 as well as ATACseq profiles between SEM or SEMPINO_RES, suggesting that acquired pinometostat resistance does not lead to obvious global changes in open chromatin (Fig. 2A). Upon analyzing gene expression profiles, it became apparent that in the absence of pinometostat, there was a noteworthy decrease in the expression of 760 out of the 13,371 genes expressed (5.7%), while 588 genes (4.4%) exhibited an increase in expression in SEMPINO_RES cells in comparison to SEM cells (Fig. 2B, Table S1). The differences in gene expression patterns triggered by pinometostat were relatively less prominent between the two cell line models (Figure S2, Table S1). In the presence of pinometostat in the original SEM cells, 670 genes (5.0%) revealed a significant decrease in expression, and 596 genes (4.5%) demonstrated a notable increase in expression, when compared to untreated SEM cells. Conversely, in the presence of pinometostat in SEMPINO_RES cells, 208 genes (1.6%) were significantly downregulated, while 388 genes (2.9%) were significantly upregulated compared to untreated SEMPINO_RES cells. Interestingly, a considerable number of genes reported to represent potential target genes of KMT2A fusion proteins [9, 20, 24, 25] were significantly downregulated in SEMPINO_RES cells (Fig. 2C, Table S1). Approximately half of the KMT2A::AFF1 target genes identified by Guenther et al. [24] were downregulated in SEMPINO_RES (Fig. 2D, Table S1), as well as a quarter of the top 50 genes associated with H3K79 methylation in KMT2A-rearranged acute leukemia patient samples as identified by Krivtsov et al. [9].
Fig. 2
Characterization of SEMPINORES by RNA-, ATAC-, and ChIP-sequencing. A. Heatmap showing ChIP-seq reads of KMT2A, AFF1, H3K79me2, and ATAC-seq reads at all KMT2A::AFF1 binding sites in SEM cells as well as SEMPINO_RES at the same location, ranked by peak width. Scale bar represents normalized read count. B. Pie chart showing the number of genes for which the expression was significantly (i.e., at false discovery rate (FDR) adjusted p-values of < 0.05) downregulated (blue), upregulated (red), or remain unchanged (gray) between SEMPINO_RES in comparison to SEM (RNA-seq data; n = 4 biological replicates/sample). C. Venn diagram showing the overlap of downregulated (blue) or upregulated (red) putative KMT2A-fusions target genes (n = 181) (white) in SEMPINO_RES compared to SEM. The putative KMT2A fusion target genes in this figure comprise the combination of genes identified by four independent studies [9, 20, 24, 25], and D. similar Venn diagrams are presented using the KMT2A fusion target genes from each individual study. E. Forest plot showing hallmark gene sets that were positively or negatively enriched in Geneset Enrichment Analysis (GSEA), based on the Normalized Enrichment Score (NES). F. Heatmap showing the most positively enriched and significantly upregulated genes (n = 50) as well as the most negatively enriched and significsntly downregulated genes (n = 50) in SEMPINO_RES as determined by GSEA. Data shown represents normalized RNA-seq counts in SEM and SEMPINO_RES cells cultured for 7 days in either the absence (-) or presence (+) of 50 µM pinometostat of n = 4 biological replicates
Likewise, ~ 25% of the genes we previously reported to be differentially expressed in response to siRNA-mediated repression of KMT2A::AFF1 and KMT2A:MLLT1 in KMT2A-rearranged ALL cells, [20] as well as a fourth of the genes reported to display binding of KMT2A:AFF1 that spreads beyond the gene promoter and well into the gene body as recently identified in SEM cells by Kerry et al., [25] were downregulated in SEMPINO_RES (Fig. 2D, Table S1). Thus, acquired resistance to DOT1L inhibition leads to selective (or partial) loss of KMT2A-fusion driven gene expression. To explore biological pathways potentially affected by acquired resistance to DOT1L inhibition, we performed Gene Set Enrichment Analysis (GSEA) on all RNA-seq data, and identified various hallmark gene sets to be significantly (nominal p-value < 0.05) modulated in SEMPINO_RES. These included the upregulated gene sets ‘MYC targets v1 and v2’ and ‘Oxidative Phosphorylation’, as well as downregulated gene sets such as ‘p53 pathway’ (i.e., DNA damage response genes), ‘epithelial-to-mesenchymal transition (EMT)’ and ‘hypoxia’ (Fig. 2E).
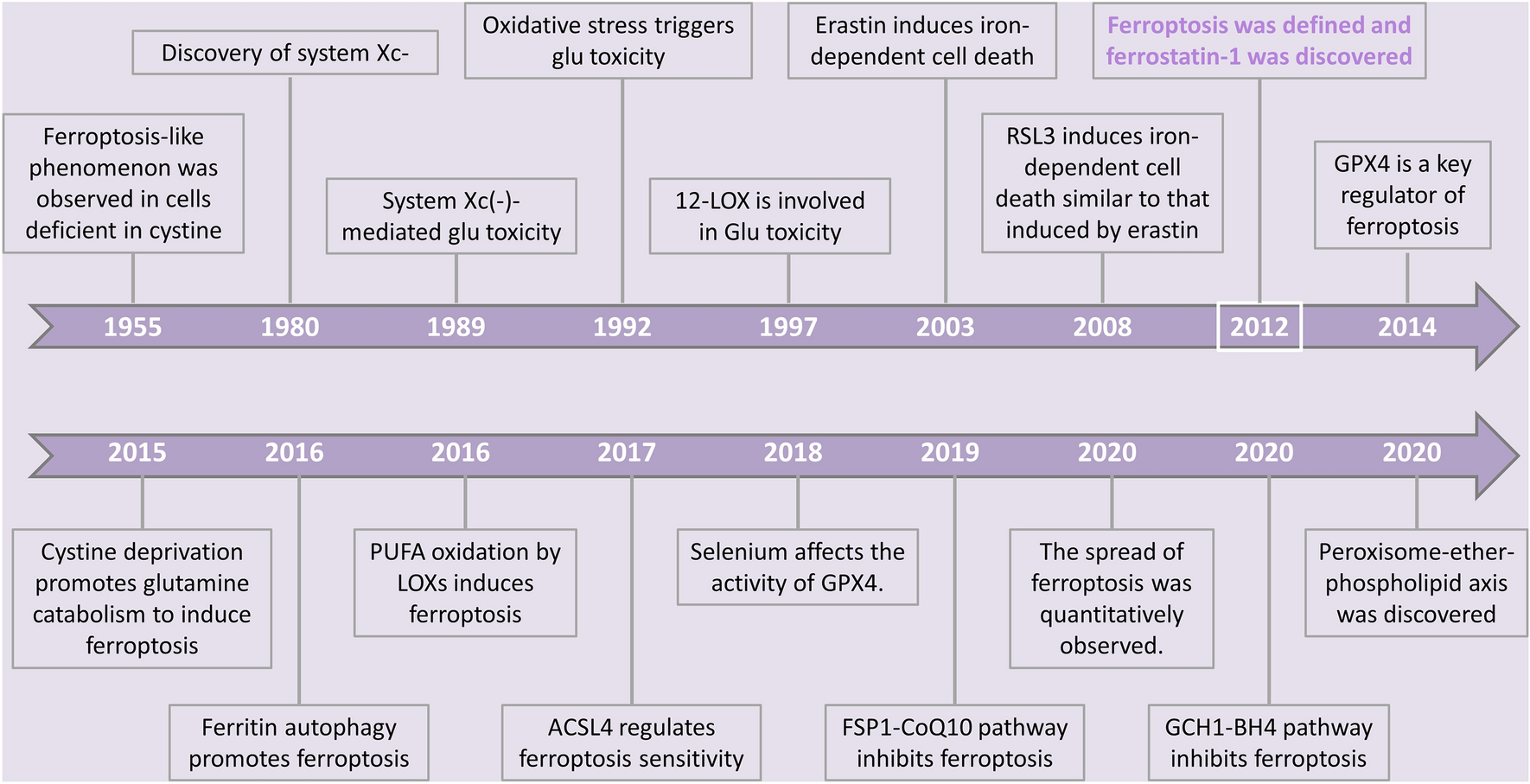
Examination of the genes most prominently enriched (n = 50) and the genes most notably under-represented (n = 50) in our GSEA data revealed PROM1 to be the most downregulated gene (GSEA score of -4,83) and CCNA1 the most positively enriched gene (GSEA score of 3,66) in SEMPINO_RES cells (Fig. 2F). Both genes represent putative KMT2A fusion targets epigenetically marked by H3K79 methylation and have shown to be highly and specifically expressed in KMT2A-rearranged ALL [9]. PROM1 encodes a transmembrane glycoprotein (i.e., CD133) commonly regarded as a cancer stem cell marker [26,27,28] and reported to be an important target of KMT2A::AFF1 [24, 29, 30]. PROM1 is robustly expressed in SEM cells but readily downregulated during pinometostat exposure, whereas PROM1 expression was nearly absent in SEMPINO_RES (Fig. 3A). Analysis at protein level by immunoblot and FACS confirmed the complete loss of PROM1/CD133 in SEMPINO_RES, while in SEMPINO_INT PROM1/CD133 was still present in in ~ 88% of the cells (Fig. 3B-E). This indicates that PROM1/CD133 expression is gradually lost from the population after prolonged pinometostat exposure. Similarly, RS4;11 cells firmly express PROM1/CD133, which was markedly reduced in both RS4;11PINO_RES#1 and RS4;11PINO_RES#2 (Fig. 3B-E).
Fig. 3
Acquired resistance to DOT1L inhibition leads to selective loss of KMT2A-fusion driven PROM1 expression. A.PROM1 mRNA expression in SEM and SEMPINO_RES cells cultured for 7 days in the absence (-) or presence (+) of 50 µM pinometostat, as determined by RNA-seq. Values indiacte normalized counts with SD derived from four biological replicates for each cell line and condition. ****p < 0.0001. B. Western blot images of PROM1 and GAPDH protein levels in indicated cell line models, and C. quantification of PROM1 expression relative to GAPDH by densitometry analysis. Values indicate mean ± SD PROM1 protein expression as determined in two biological replicates. *p < 0.05. D. Histograms showing the counts of viable cells positive for PROM1/CD133 of indicated cell line models, as determined by flow cytometry (FACS) analysis, and E. Quantification of PROM1/CD133 expression presented as the mean ± SD as determined by two independent FACS experiments. *p < 0.05, **p < 0.005. F. Differences in chomatin accessibility at the PROM1 and TAPT1 gene locus between SEMPINO_RES and SEM cells as determined by ATAC-sequencing of two biological replicates (on top). Vertical blue lines indicate significant decreases of chromatin accessibility in SEMPINO_RES cells, whereas grey lines indicate equal chromatin accessibility in both SEMPINO_RES and SEM. The ATAC-sequencing results are followed by ChIP-sequencing tracks of the same locus showing the distribution of H3K79Me2, H3K27Ac, H3K4Me3, KMT2A in SEM and SEMPINO_RES cells cultered for 7 days in either the absence (-) or presence (+) of 50 µM pinometostat.
Differences were statistically evaluated using unpaired t-tests
ChIP-sequencing data for KMT2A, AFF1, H3K4me3, H3K79me2, and H3K27ac showd that upon pinometostat exposure, SEM cells display a clear reduction of KMT2A binding in PROM1, which was accompanied by strong reductions of the levels of H3K79me2, H3K4me3, and H3K27ac at the PROM1 gene as well as at its enhancer TAPT1 [29] (Fig. 3F). In untreated SEMPINO−RES cells the PROM1 locus is completely devoid of KMT2A, AFF1, H3K79me2, H3K4me3, and H3K27ac, suggesting that this gene is no longer being regulated by KMT2A::AFF1 and subsequent DOT1L-mediated H3K79 methylation. Also, ATAC-sequencing clearly revealed decreased chromatin accessibility at the promoter and enhancer of PROM1 in SEMPINO−RES (Fig. 3F). Interestingly, at the TAPT1 locus in these same SEMPINO−RES cells KMT2A and AFF1 binding as well as the levels of H3K4me3, H3K79me2, and H3K27ac to some extent remained intact (Fig. 3F).
In addition to PROM1, the expression of other putative KMT2A::AFF1 target genes, including RUNX2, PRSS12, ZC3H12, SERPINB1, GNAQ and BANK1 were severely downregulated in SEMPINO−RES with a logFC of > 3-fold (Fig. 2F and Figure S3A) and exhibited similar patterns of RNA-, ChIP-, and ATAC-seq as observed for PROM1 (Figure S3B), indicating their dependence on KMT2A::AFF1-mediated epigenetic control. In contrast, at SERPINB1 only moderate levels of H3K79me2 were observed, accompanied by rather weak binding of KMT2A and absence of AFF1, suggesting that this gene may not necessarily be regulated KMT2A fusion proteins and DOT1L (Figure S3A, B),).
Collectively, these data demonstrate that a selection of known KMT2A::AFF1 target genes that are responsive to pinometostat-mediated DOT1L inhibition in SEM cells are relieved from the epigenetic control of KMT2A::AFF1 and become transcriptionally silenced in SEMPINO_RES cells.
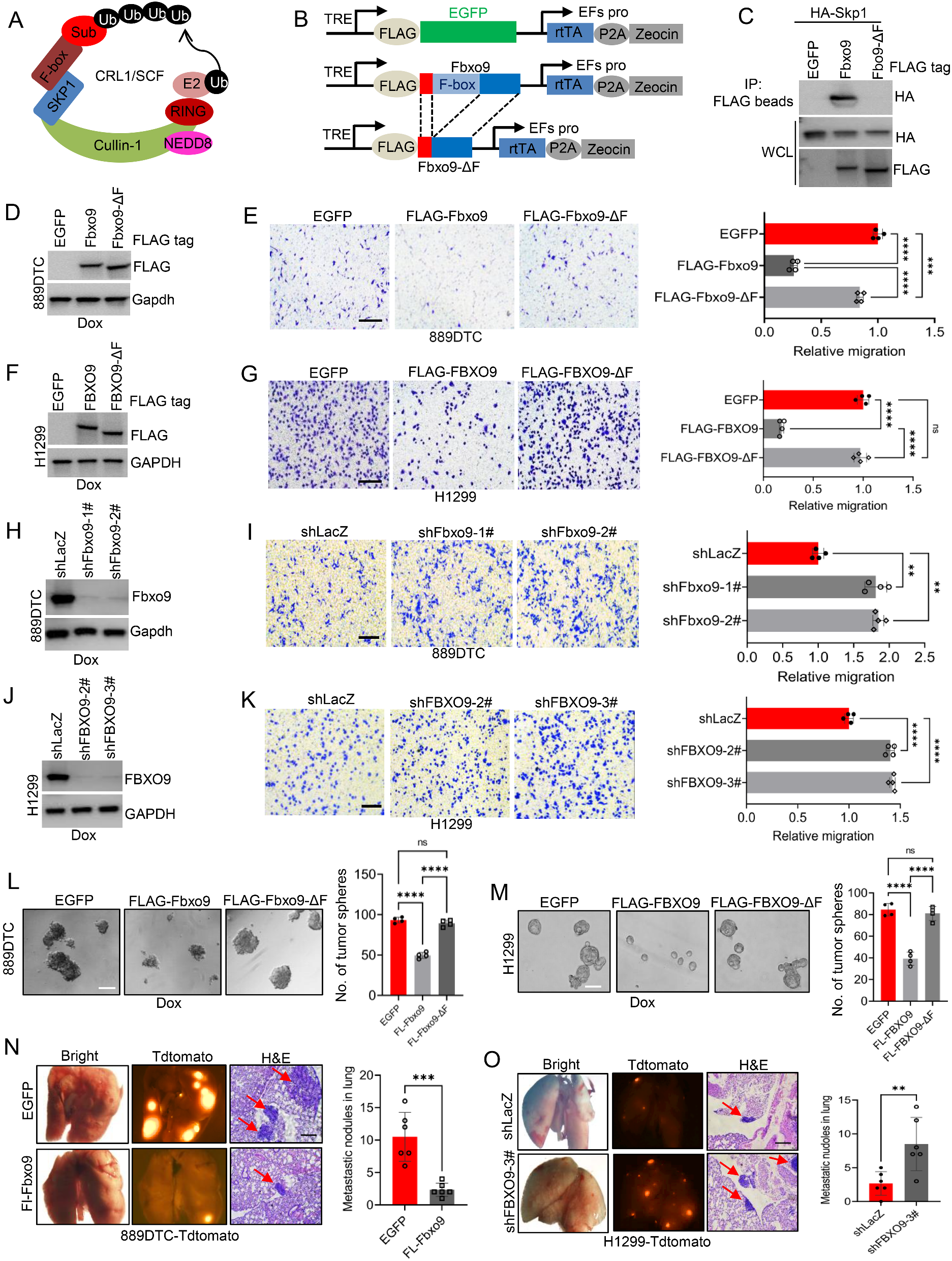
Following previous evidence on the role of DOT1L in HOXA gene expression in KMT2A::AFF1+ ALL cells, [9, 24] we examined the HOXA locus and found that HOXA9 and HOXA10 were expression at comparable levels in both SEM and SEMPINO_RES (Fig. 4A). Inhibition of DOT1L-mediated H3K79 methylation by pinometostat resulted in moderately decreased expression of HOXA9, HOXA7, and HOXA10, while the levels of H3K27ac, H3K4me3, KMT2A, and AFF1 remained unchanged in both cell lines (Fig. 4A,B). Similar patterns were found for other KMT2A-fusion target genes, including CDK6, involved in cell proliferation in KMT2A rearranged ALL [20] (Fig. 4A,B and S4A-C). The expression of MEIS1, which encodes a required co-factor of HOXA9-driven leukemogenesis, [9, 24, 31, 32] remained unaltered upon pinometostat exposure despite reductions in the levels of H3K79me2 in both SEM and SEMPINO_RES (Fig. 4A,B and S4A,B,C). Collectively, this indicates that a subset of KMT2A-fusion target genes continued to be regulated by DOT1L in pinometostat-resistant ALL cells, while a separate group of genes showed no transcriptional response to the inhibition of DOT1L-mediated H3K79 methylation.
Fig. 4
Unaltered or upregulated gene expression of KMT2A-fusion targets after acquired resistance to DOT1L inhibition. A. HOXA7, HOXA9, HOXA10, MEIS1, CKD6, and CCNA1 mRNA expression SEM and SEMPINO_RES cells cultured for 7 days in the absence (-) or presence (+) of 50 µM pinometostat, as determined by RNA-seq. Values indiacte normalized counts with standard deviation (SD) derived from four biological replicates for each cell line and condition. Differences in expression were statistically evaluated using unpaired t-tests; * p < 0.05, * p < 0.05, ** p < 0.005, *** p < 0.0005, **** p < 0.0001. Differences in chomatin accessibility at the HOXA, MEIS1, CKD6, and CCNA1 gene loci between SEMPINO_RES and SEM cells as determined by ATAC-sequencing by two biological replicates (on top). Vertical blue lines indicate significant decreases of chromatin accessibility in SEMPINO_RES cells, whereas grey lines indicate equal chromatin accessibility in both SEMPINO_RES and SEM. Red lines indicate significant increases in chromatin accessibility in SEMPINO_RES. Below the ATAC-sequencing data, ChIPseq tracks showning the presence of H3K79Me2, H3K27Ac, H3K4Me3, KMT2A, and AFF1 at the corresponding gene loci in SEM and SEMPINO_RES cells cultured for 7 days in the abscence (-) or presence (+) of 50 µM pinometostat
Intriguingly, we also found the expression of some putative KMT2A-fusion target genes to be upregulated in SEMPINO_RES in the absence of pinometostat (Figure S4D), including HOXA7, NLGN4X, CCNA1, FCRLA, IL7R, LYN and FUT4 (Fig. 4A,B and Figure S4D,E).
Upregulation of myeloid-associated gene expression in KMT2A-rearranged ALL cells upon acquired resistance to DOT1L inhibitionApart from differential gene expression of putative KMT2A::AFF1 target genes, our data also revealed changes in expression of genes not associated with KMT2A fusions and/or H3K79 methylation (Fig. 2F). One of the most enriched and upregulated genes upon acquired pinometostat-resistance according to our GSEA is LILRB4 (Figs. 2F and 5A), encoding the monocytic differentiation marker CD85k [33,34,35]. LILRB4 is hardly expressed in SEM cells but is moderately upregulated during pinometostat exposure and substantially expressed in SEMPINO_RES cells (Fig. 5A). In SEM cells, pinometostat induced an increase of chromatin accessibility as well as an increase in the levels of H3K27ac and KMT2A binding at the LILRB4 locus, yet no H3K79me2 or binding of AFF1 was detected, suggesting that upregulated of LILRB4 expression is not dependent on DOT1L or KMT2A::AFF1 (Fig. 5B).
Fig. 5
Upregulation of myeloid-associated CD85k/LILRB4 and CD33 expression during the acquirement of resistance to DOT1L inhibition. ALILRB4 mRNA expression in SEM and SEMPINO_RES cells cultured for 7 days in the absence (-) or presence (+) of 50 µM pinometostat, as determined by RNA-seq. Values indiacte normalized counts with SD derived from four biological replicates for each cell line and condition. Differences in expression were statistically evaluated using unpaired t-tests; **p < 0.005, ***p < 0.0005. B. Differences in chomatin accessibility at the LILRB4 gene locus between SEMPINO_RES and SEM cells as determined by ATAC-sequencing by 2 biological replicates (on top). Red boxes indicate locations within the LILRB4 gene locus of significantly increased of chromatin accessibility in SEMPINO_RES as compared to SEM cells. In addtionm, ChIPseq tracks are presented showing the presence of H3K79Me2, H3K27Ac, H3K4Me3, KMT2A, and AFF1 at the same locus in and indicated cell line models cultured for 7 days in the absence (-) or presence (+) of 50 µM pinometostat. C. Histograms showing the counts of viable cells positive for CD85k/LILRB4 and D. CD33 protein surface expression of indicated cell line models, as determined by flow cytometry (FACS) analysis. Fluorescence Minus One (FMO) controls were used to determine the cut-off point for the positive cell population. E. Quantification of CD85k/LILRB4 and F. CD33 expression represented as the mean ± SD, determined through either one (RS4;11 cells) or two (SEM cells) independent FACS experiments, each involving biological replicates. Differences in expression were statistically evaluated using unpaired t-tests; *p < 0.05, **p < 0.005, ***p < 0.0005
FACS analysis confirmed an increased population of ~ 60% in LILRB4/CD85k positive cells in SEMPINO_RES compared to only ~ 7% in SEM (Fig. 5C,E). Counterintuitively, instead of an expected moderate increase in LILRB4/CD85k-positive cells, we found SEMPINO_INT to have lost LILRB4/CD85k expression almost completely (Fig. 5C,E). In RS4;11 already 25% of the cells were positive for LILRB4/CD85k, which tremendously increased to approximately 90% of the cells in both RS4;11PINO_RES daughter lines (Fig. 5C,E). Interestingly, apart from LILRB4/CD85k, we found additional myeloid-associated genes to be upregulated in SEMPINO_RES, including CD33, CCL5, LIMK1, and MPEG1, revealing similar patterns of RNA-, ChIP- and ATAC-seq as LILRB4, although less prominent (Fig. 2F, Figures S5A, S5B). CD33, commonly expressed in a subpopulation in KMT2A-rearranged infant ALL [36,37,38], serves as an important immunophenotypic marker for the characterization of pediatric acute myeloid leukemia (AML) by EuroFlow [39,40,41,42] and has been exploited as a therapeutic target for AML. In SEM a subpopulation of 20% of CD33-positive cells was identified, which was increased in SEMPINO_RES to about 40%, yet CD33-positive cells again were largely absent in SEMPINO−INT (Fig. 5D,F). Similarly, in RS4;11 a CD33-positive subpopulation of 9% was increased upon pinometostat resistance to 14% in RS4;11PINO_RES#1 and up to 50% in RS4;11PINO_RES#2 (Fig. 5D,F).
Together these data indicate that under prolonged pressure of DOT1L inhibition, KMT2A-rearranged ALL cells seem to initiate a reprogramming process that involves the acquisition (or selection) of myeloid-like characteristics.
Drug screens reveal minimal cross resistance, and sensitization towards venetoclax after acquired pinometostat resistanceFinally, acquired pinometostat resistance led to the upregulation of the multidrug efflux pump ABCB1 (Fig. 2F and Figure S6A,B), associated with multidrug resistance and previously reported as the mechanism of pinometostat resistance in KMT2A-rearranged acute leukemia cell lines [17]. However, our data challenges the concept that elevated ABCB1 expression alone is the mechanism of resistance to DOT1L inhibition as reported previously [17]. Despite significant ABCB1 upregulation, we still observe comparable inhibition of H3K79 methylation in SEM and SEMPINO_RES (Fig. 1D and E), indicating that the amount of pinometostat and/or its retention in SEMPINO_RES cells is sufficient to effectively reduce H3K79me2 levels, overriding the impact of ABCB1 upregulation.
Since multidrug efflux pumps are associated with multidrug resistance [43, 44], we assessed whether SEMPINO_RES cells had become more resistant to current chemotherapeutics for KMT2A-rearranged infant ALL [1, 2] and whether we could identify agents to which SEMPINO_RES cells had become more sensitive by performing drug library screens (Table S2, Fig. 6A,B).
Fig. 6
Moderate levels of cross-resistance and substantial sensitization towards venetoclax in pinometostat-resistant KMT2A-rearranged ALL cells. A. Drug response curves for prednisolone, dexamethasone, vincristine, daunorubicin, cladribine, cytarabine, and L-asparaginase as determined by 4-day MTT assays in SEM and SEMPINO_RES cells with n = 4 biological replicates, each comprising n = 3 technical replicates, and presented as the mean ± standard error of the mean (SEM). B. Comparison of the IC50 in SEM and SEMPINO_RES cells for a total of 679 compounds tested in dru
留言 (0)