1. IntroductionIlheus virus (ILHV), a neglected human pathogen, is believed to be maintained in nature by an enzootic cycle between mosquito vectors and birds [
1]. Structurally, ILHV has a spherical morphology with enveloped positive-sense single-stranded RNA and belongs to the genus Flavivirus in the Ntaya antigenic serocomplex [
2,
3]. It is phylogenetically associated with the Rocio virus (ROCV) with 73.37–74.5% amino acid identity, and ILHV and ROCV are considered members of the same species [
4]. ILHV infection in humans can lead to Ilheus fever, an acute undifferentiated febrile disease with non-outbreak cases. However, the clinical spectrum can vary in severity, and the fever can progress to severe life-threatening meningoencephalitis [
5,
6,
7,
8,
9,
10]. In 2017, the first documented case of a fatality following ILHV detection via nucleic acid amplification in an elderly patient with encephalitis in Brazil drew public attention to the need to better understand the clinical–epidemiological scenario [
11]. It is important to mention that in this case, it was not determined whether the virus caused encephalitis in the patient, who had co-morbidity (diabetes and hypertension).Historically, ILHV was isolated in 1944 from a pool of Psorophora and Aedes mosquito species captured near Ilhéus city in Bahia state, Brazil, during an entomological surveillance of the yellow fever virus vector [
1]. Viral infection in humans occurs via mosquito bites, especially from mosquitoes of the genera Psorophora and Ochlerotatus. ILHV was isolated from humans in 1959 in Belém city (Pará, Brazil). This was followed by viral isolation in Trinidad, West Indies (1962), Colombia (1966), Panama (1967), French Guiana (1973), Brazil (1995), and Ecuador (2007) [
5,
7,
8,
9,
12,
13,
14]. Currently, several arboviral surveillance studies, including those on ILHV isolates, molecular identification, and serosurvey to detect antibodies against ILHV, have been carried out in the last few decades in South and Central America and in the Caribbean islands. Thus, according to the results of these studies, ILHV infections have been observed in several vertebrate species, such as monkeys, birds, rodents, bats, horses, coatis, and buffaloes (
Supplementary Files S1 and S2) [
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33,
34,
35,
36].
However, several barriers related to the difficulty in ILHV detection remain. The main factors include diverse endemic mosquito-borne flavivirus populations, a short viremic period, lack of specific clinical features, and the possibility of serological tests exhibiting cross-reactivity to shared epitopes on other endemic flaviviruses. Overall, these factors contribute to the gap in estimating the true ILHV burden on the population.
A clear and comprehensive understanding of the overall exposure rates to ILHV and its clinical manifestations is necessary to provide useful information for healthcare workers and policymakers and to underpin effective prevention and control strategies. However, these studies are lacking in scientific literature. The aims of this study were to compute the pooled rate of exposure to laboratory-confirmed cases of ILHV infection and describe the clinical characteristics of individuals following ILHV infection.
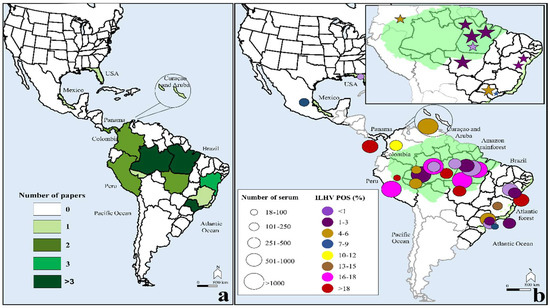
4. DiscussionTo the best of our knowledge, this is the first meta-analysis of ILHV infection. The main finding of the rates of ILHV seropositivity (9%, 95% CI: 5–13) included 17,722 individuals, with serological samples collected over many decades (1958–2015). Even when considering only the monotypic reactions, positivity remained at a considerable rate of 2% (95% CI: 1–2), with an indication of ILHV prevalence beyond the Amazon region (including parts of Brazil, Colombia, and Peru), as there was also positivity in the Brazilian states of Bahia and São Paulo. The moderate prevalence of ILHV-neutralizing antibodies, coupled with the low number of clinical cases of ILHV infection, suggests that most infections are inapparent or mild, which may be an important vehicle for viral spread to non-endemic regions [
2,
72,
73].Through our meta-analysis, we were able to elucidate the clinical characteristics of ILHV infection. Fever, headache, myalgia, encephalitis, malaise, arthralgia, gastrointestinal distress, and cough were the most frequently described clinical features. Since the infection is indistinguishable from other febrile syndromes and has a wide spectrum of clinical severity, ranging from subclinical to severe, a portion of symptomatic ILHV infections could be masked by clinical manifestations associated with multiple human pathogenic arboviruses [
74], which may contribute to the underestimation of the burden of ILHV. Notably, no case of long-term sequelae or death has been linked to ILHV infection. This fact demands continuous efforts to establish effective public health arbovirus surveillance programs because although the number of confirmed cases of the disease is low, there is a high probability of underreporting due to difficulties in accurate laboratory diagnosis.Regarding
Figure 4, for the two studies from 2014–2015 [
70,
71], it is difficult to explain this high rate of seropositivity (mean = 58%, 95% CI: 55–62), but it is interesting to mention that these studies involved populations at high risk for arboviruses: male military personnel of working age in the Amazon rainforest or rural population. A high rate of heterotypic serological reactions were observed. These variables were also present in most of the other studies included in the meta-analysis; however, except for Buckley et al. [
54], all other studies had lower ILHV positivity.In South America, Central America, and the Caribbean, different geographic regions have multiple endemic mosquito-borne flaviviruses in co-circulation [
75,
76,
77]. Twenty-nine (78.3%) of the 37 studies included in our meta-analysis investigated the antibody ratios against multiple arbovirus antigens. Flavivirus species have been linked to sporadic cases of human infection, including Bussuquara (BUSV), Cacipacore (CPCV), and ILHV [
78,
79]. All these viral pathogens may cause undifferentiated febrile illness in humans, and laboratory diagnosis is difficult due to the substantial cross-reactivity of antibodies [
75]. Additionally, as occurs for ILHV, BUSV, CPCV, and other native flaviviruses are neglected, with misdiagnosis of cases compared to other medical arboviruses associated with outbreaks or epidemics that attract the most public attention, such as DENV, ZIKV, and YFV.ILHV shares antigenic similarities with Bagaza, Ntaya, and Tembusu viruses in cross-neutralization tests [
80]. These antigenic similarities are attributed to their genetic similarities. Interestingly, serocomplex cross-reactive memory can elicit viral cross-protection against infections with different serocomplexes [
81,
82]. In other words, polyclonal antisera (post-immune) against a specific virus can neutralize other viruses. As a result, Ilheus and Saint Louis encephalitis (SLE) flavivirus infections engender cross-protection against a lethal ROCV challenge in mice [
83]. Similar cross-protection has also been demonstrated between Chikungunya and Mayaro alphaviruses and between DENV and West Nile and SLE flaviviruses [
84,
85]. New insights into protective ILHV infection responses are needed to elucidate its effects on previous flavivirus exposure and subsequent ILHV exposure. In addition, it is also important to establish whether ILHV immunity provides certain types of cross-protection against other co-circulating flaviviruses in certain geographic locations. Clarification of these factors is essential for a better understanding of the epidemiology of ILHV infection and for improving diagnostic methods.Although seroepidemiological surveys are useful, this method could overestimate the frequency of ILHV positivity owing to cross-reactivity with other flavivirus infections. Since the included studies used HI/NT tests to capture total antibodies in the target serum, it was not possible to determine the dominant ILHV antigen (protein) involved. There is marked variation in the amino acid identity of the polyprotein of mosquito-borne flaviviruses; for example, in the polyprotein of Japanese encephalitis, dengue, yellow fever, Zika, and West Nile viruses, the amino acid similarity ranges between 45–77% [
75]. As HI and serum neutralization tests target neutralizing antibodies, virus E (envelope) protein may be a promising target. The E protein shares 43–79% amino acid similarity between flaviviruses. The most cross-reactive ILHV antigen is the NS3 protein, which shares amino acid homology with the JEV complex [
34]. In our meta-analysis, the cross-reactivity of serodiagnostic tests was likely modulated because we were able to demonstrate data on monotypic-type reactions [
51,
52,
56,
59,
60,
61,
63,
67,
69,
71].The HI assay has been a mainstay serodiagnostic test for a long period of time, with the first assay described in the 1940s used to measure the hemagglutinating activity of the influenza virus [
86]. Toward the end of the 1950s, antigens prepared from the Laemmert strain of ILHV were used in the HI assay [
46]. The HI assay is highly applicable for the detection of anti-ILHV antibodies because no licensed kits are available for this purpose, and in-house HI presents low equipment and laboratory supply costs. A potential drawback is that this method is not commonly standardized, as is the plate-reduction neutralization test (PRNT) [
87]. Thus, it is necessary to empirically determine how to weigh antibody cross-reactivity when interpreting the results. Therefore, new insights into ILHV serology assays that aid in determining the specificity (antibody cross-reactivity rate) and sensitivity across current and future laboratory techniques are of paramount importance.A key factor to consider is the lack of detailed analyses of cross-neutralization studies involving ILHV cases; such studies may contribute to the level of uncertainty in ILHV diagnoses. This potential uncertainty may be mitigated by the studies included in our analysis (more than one-third) that used the viral neutralization assay, that is, NT, and showed good specificity [
36,
46,
48,
51,
54,
57,
63,
65,
66,
88,
89]. Herein, in direct comparisons of NT and HI, similar results were noted, with overall ILHV positivities of 7% and 9%, respectively. Certainly, the search for neutralizing antibodies by PRNT, considered the “gold standard” in discriminating serodiagnosis of flavivirus infections, will contribute in reducing false-positive cases. However, PRNT may require handling live pathogens in a biosafety level 3–4 laboratory and is a labor-intensive and costly technique for routine laboratory use, making it difficult to be employed on a large scale [
88,
89].
Our study has important strengths, including the meta-analytic approach and the inclusion of 37 studies describing the serological and clinical characteristics of a large number of individuals. Although studies with small sample sizes were included, they did not impact the estimated pooled rate of exposure, likely because only three studies had sample sizes <100 individuals. The potential limitations of our study include the inconsistent availability of data in some of the selected studies. Several studies failed to report basic demographic details, such as sex, age, and professional activities, and were therefore excluded from subgroup analyses. Demographic variables are important as they enable the assessment of potential risk factors associated with higher seroprevalence. Further studies with complete data are required. Second, because most studies were not designed to determine the prevalence and incidence rates of ILHV infection, it was only possible to estimate the individual and combined proportions regarding the frequency of positivity of the pathogen. Therefore, future studies that are better distributed in the Americas and have data on monotypic serological reactions are necessary. A third potential limitation was the heterogeneity observed in the majority of the analyses, which was possibly due to differences in epidemiological settings, such as the endemicity of place linked with transmission intensity, differences in population groups with greater or lesser probability of exposure to the mosquito vector, laboratory accuracy of the techniques, and varied period of time.


Comments (0)