Participants
Thirty healthy adults participated in the study (17 women and 13 men) aged 19–40 years (mean age: 29.8years, SD: 6.6 years). The choice for the number of participants was based on an a priori power analysis using G*power (Version 3.1.9.6). The design was a single group repeated measures ANOVA with 3 repeated measurements (corresponding to diurnal variation or repetitions). Effect size was set to 0.25 (medium), alpha probability was set to 0.05 and power was set to 0.8. Using these criteria the estimated participant number was 28, with the actual n of 30 giving a power of 0.84. Participants had an average 16.9 years of education (SD: 2.6years, range: 12–23 years). Exclusion criteria for all participants included the presence of any neurological or psychiatric disorder and the use of psychotropic or antihistamine medication. All participants were instructed to maintain their usual level of activity and to have a subjectively satisfactory level of sleep on the days on which the experiment was conducted.
The study protocol was approved by the Ethics Committee of the University Mental Health Research Institute. Participants were provided with written information about the purpose and duration of the study, the nature of the experimental tasks, and the personal data collected, which were treated anonymously and confidentially, in accordance with the legislation and the relevant provisions of the General Data Protection Regulation (GDPR) of the European Parliament and Council. Participants signed informed consent forms, informing them of the voluntary nature of their participation and their right to withdraw at any point during the study.
Procedure
The 30 participants were divided into three groups, each consisting of ten people. The experiment for each participant consisted of three sessions over a 24-hour period during one or two calendar days, depending on the group to which the participant was assigned. In each session the participant performed all oculomotor tasks. The timing of each session was determined by the time of morning awakening based on the relevance of this schedule to internal circadian rhythms and especially the circadian variation of cortisol with highest cortisol levels at 30 min post awakening and lowest late in the evening (Debono et al. 2009). Thus, each participant underwent a morning, afternoon, and evening session at 1-, 6-, and 12-hours post-awakening, respectively. The mean absolute time for the morning measurement for all participants was 10am (SD: 1 h ), for the afternoon measurement it was 15pm (SD: 1 h ) and for the evening measurement it was 21pm (SD: 1 h). The groups, however, differed in the time of the day they started the experiment. This resulted in three groups: the first followed the sequence morning—afternoon—evening, the second followed the sequence afternoon—evening—next morning and the third followed the sequence evening—next morning—next afternoon. One member of the research group visited the participant at his/her home three times to collect the data. Weekend days or days-off from work were selected so the participant was at home. We did not use some specific instrument for quality of sleep and physical activity measurement. However, we asked the participants to try to have a good night sleep and avoid strenuous physical activity and consuming alcohol for the day of measurement, which was a weekend day during which they remained at their home for most of the time. We relied on self-reports of participants for complying with these instructions.
Eye movement recording
Oculomotor tasks were performed using the “Saccadometer Plus” (Ober Consulting Sp. z.o.o. Poznań, Poland), a portable device, with unrestrained head movement, easily used outside of laboratory environment that detects horizontal eye movements, with sampling frequency of 1 kHz, using the technology of Direct Infra-Red Oculography. The device records infrared reflections from the medial limbus of each eye, which then are compared to produce a single measure of horizontal gaze position, under the assumption that conjugate eye movements are being made. Three visual targets are projected by the device itself by means of three laser emitting light sources located above the eyes (forehead), one centre and two peripheral ones to the right and left of the centre at equal distances of 10 degrees of visual angle. This set up produces peripheral targets at a fixed angular separation of 10 degrees from the centre target, regardless of the separation between the device and the projection surface. Measurements were performed in a quiet room under good lighting conditions with additional lighting, if needed, coming from fluorescent lights. The targets were projected on a wall at 1.5–3 m, at eye level.
Oculomotor tasks
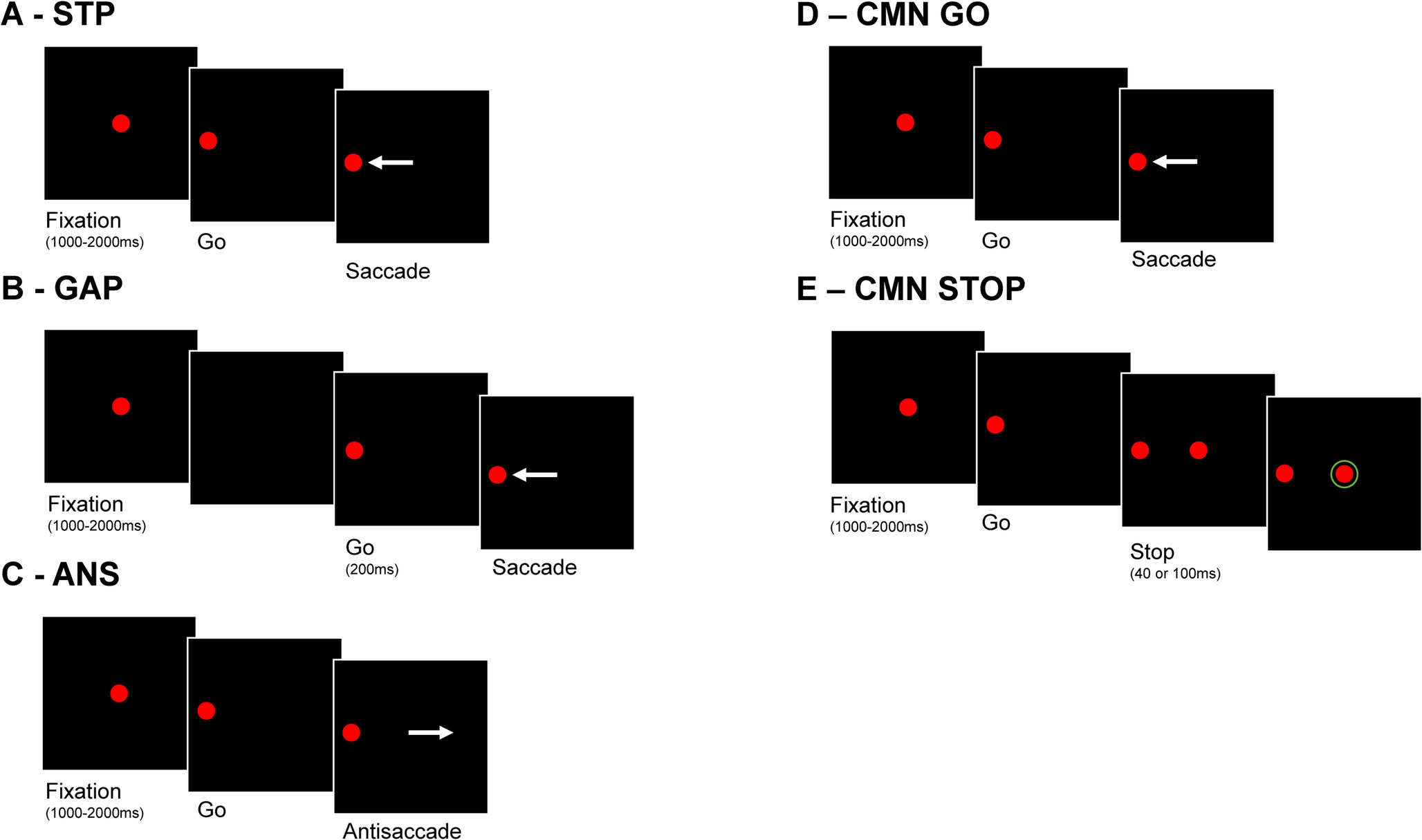
In each session four oculomotor tasks were performed by each participant: (a) the step visually guided saccade task (STP), (b) the gap-visually guided saccade task (GAP), (c) the antisaccade task (ANS) and (d) two blocks of the countermanding saccade task (CMN). The STP started with fixation on the central target for a period selected randomly at each trial between 1000 and 2000ms (uniformly distributed). Then the central target disappeared and simultaneously one of the peripheral targets appeared (selected randomly at each trial) and remained for a period of 1000ms. The subject was instructed to look at the peripheral target as soon as it appeared. The GAP task was identical to the STP task with the only difference being a time gap of 200ms between the disappearance of the central target and the appearance of the peripheral target. The ANS task was identical to the STP task, and the subject was instructed to look at the mirror location away from the peripheral target when this appeared. Finally, the CMN was identical to the STP task for go trials where the subject was instructed to look at the appearing peripheral target as quickly as possible, but to withhold their response if the central target reappeared (stop signal). The stop signal could appear either at 40ms after the appearance of the peripheral target or 100ms after the appearance of the peripheral target in two separate blocks of the CMN task (CMN40, CMN100).
Whilst it is recommended that different stop signals be interleaved within a block to prevent participants predicting the stop signal and so delaying their responses accordingly (Verbruggen et al. 2019), the embedded software on the occulometer we used did not allow this. Equally, the probability of the stop signal’s appearance was 50%, which was higher than has been recommended for countermanding tasks (Verbruggen et al. 2019). Given these methodological constraints, we do not attempt to calculate an absolute stop signal reaction time for our participants. The strategy of inhibiting go responses is the hallmark of the CMN task however, and so our study directly examines the magnitude of this inhibition (latency of go responses) and how successfully this allows successful countermanding (proportions of errors) for each CMN stop signal delay.
The order of the oculomotor tasks was pre-fixed and different for each participant and for each measurement of the same individual, based on a pseudo-random order predefined by protocol to ensure that there would be no order effect. Each task was completed in blocks of 100 trials, with short breaks between tasks. Before each task, there was a short training session including 10 practice trials to ensure that participants understood the task instructions and were able to perform it correctly. A calibration procedure was performed before the start of each task, using a sequence of twenty saccadic eye movements, ten to the left and ten to the right of the central fixation stimulus. We used the built-in calibration sequence of the device, readjusting the position of the tracker if the gain was not between 50 and 150 arbitrary units per degree, as recommended by the manufacturer. The calibration procedure and the 10 practice trials preceded every task repetition.
Data preprocessing and analysis
The oculomotor data were extracted using the “Latency Meter version 6.11.0 (Ober Consulting Sp. z.o.o. Poznań, Poland)”, which allowed us the visualization and verification of the oculomotor data.
In the STP and GAP tasks, the latency for the first saccadic eye movement after the peripheral stimulus presentation was recorded. After exclusion of latencies below 80ms we computed for each subject and each session the mean latency and the standard deviation of latency separately for the GAP and STP task. We then tested for the presence of a Gap effect in mean latency and SD of latency between the GAP and STP tasks using a paired samples t-test.
In the ANS task the percentage of correct antisaccades was measured as well as the mean latency and the standard deviation of latency for correct antisaccades. Again, we excluded latencies below 80 ms.
In the CMN task we computed separately for each block (40ms and 100ms of stop signal onset) the percentage of correct responses in the stop signal trials as well as the mean and standard deviation of the latency for the go trials. Again, we excluded latencies below 80ms. We then tested for significant stop signal delay effects (using paired sample t-tests). The effect of stop signal delay for each subject was defined as the difference between blocks (40ms and 100ms stop signal delay) for each CMN parameter (percentage of correct responses for the stop signal trials, mean latency and standard deviation of latency for the go trials). The longer the delay for the stop signal in the CMN task, the more difficult it is to inhibit the saccade response.
For each index of saccadic task performance as described above we performed a one-way repeated measures ANOVA to examine the effect of diurnal variation using time of day as the independent repeated measures factor (morning, afternoon, evening). The Greenhouse-Geisser correction was used to correct p values for violations of the sphericity assumption. We also performed a Bayesian repeated measures ANOVA to compute the Bayes factor, BF(1,0) favoring the alternative hypothesis that diurnal variation has a significant effect on the specific parameter and its inverse BF(0,1) favoring the null hypothesis that diurnal variation has no effect on the specific parameter. The same analyses were performed to examine the effect of repetition (first, second third) as repeated measures factor on all saccade parameters. For the ANOVA and Bayesian ANOVA analyses we used the JASP 0.19.3 software.
To test for interaction effects between diurnal variation and repetition on oculomotor function indexes we used the linear mixed effects model analysis using JAMOVI (the jamovi project 2024, version 2.6). This analysis, in contrast to the standard ANOVA analysis, allows for incomplete data sets to be used. This was the case for our data set since we could not have all combinations of repetition and diurnal variation for each subject to allow for a two factor repeated measures ANOVA (with repetition and diurnal variation as the within subject repeated measures) to be used. The linear mixed effects model analysis with random intercept was used with participant as the random intercept factor and diurnal variation and repetition as the independent fixed effect factors. We report the F values for the main effect of diurnal variation, main effect of repetition and the interaction effect.




Comments (0)