
Remember me


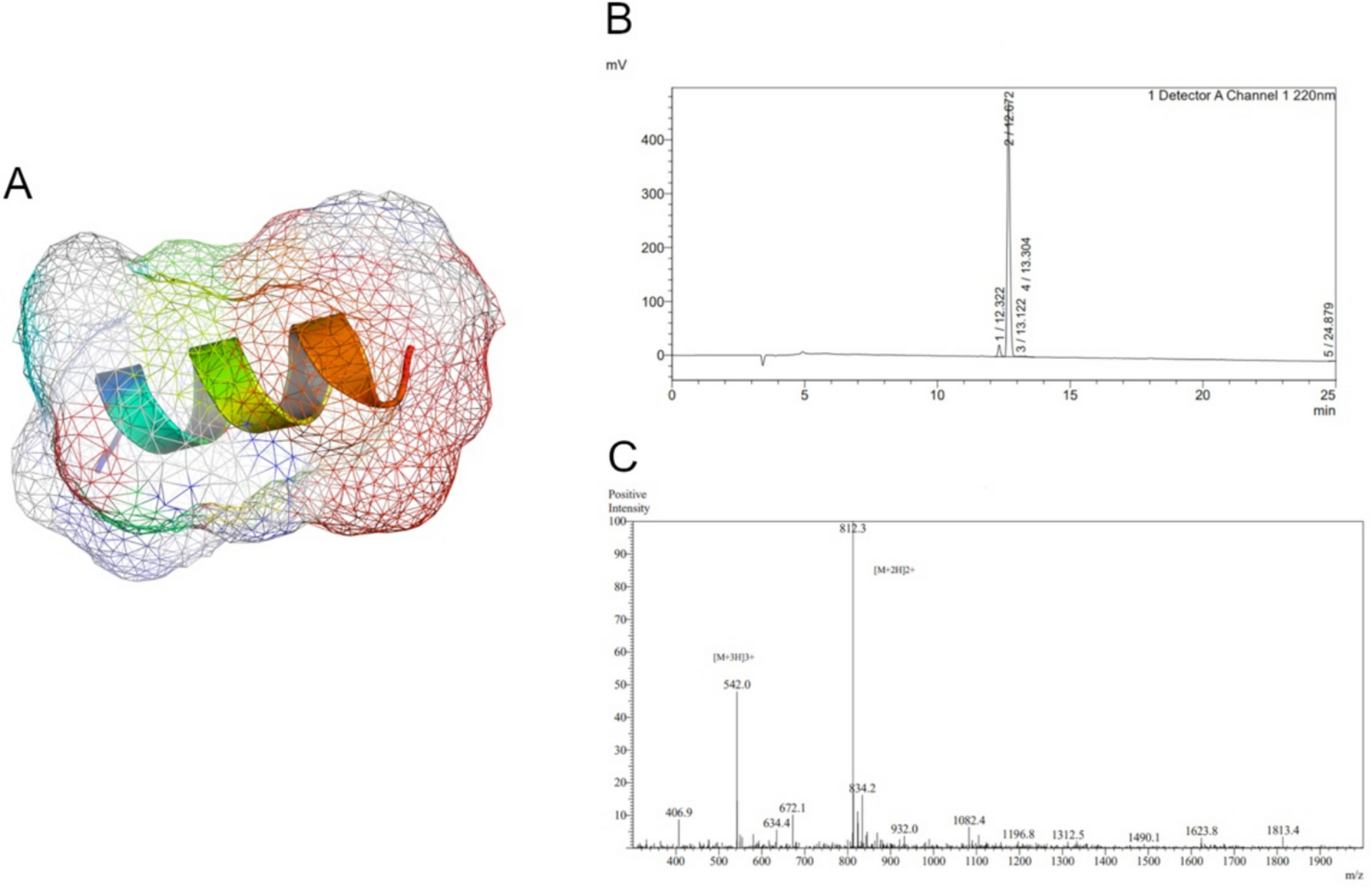



PS77, a 13-amino acid peptide (molecular weight = 1622.92 Da; Supplementary Table 1), was chemically synthesized using solid-phase peptide synthesis (SPPS). The peptide's α-helical structure, which is a common secondary structure in peptides and proteins, was confirmed (Fig. 1A). The α-helix is stabilized by hydrogen bonds between the backbone amino and carboxyl groups, and it is often associated with biological activity and stability. The presence of an α-helical structure suggests that PS77 may have a specific function and could potentially interact with target molecules or receptors. High-performance liquid chromatography (HPLC) revealed a single, sharp peak with a high intensity and with the purity of 95.3%, indicating that the PS77 peptide is highly pure and homogeneous (Fig. 1B). This high purity is essential for accurate characterization of the peptide’s properties and biological effects. Mass spectrometry (MS) analysis verified molecular weight (1622.92 Da) of PS77 (Fig. 1C). The absence of other significant peaks indicates that the peptide has a well-defined molecular mass and is not degraded or modified. The MS data confirms the identity and integrity of the PS77 peptide. Taken all together, the combination of 3D structure prediction, HPLC, and MS analyses provides a comprehensive characterization of the PS77 peptide. The predicted α-helical conformation suggests a potential biological function, while the HPLC and MS data confirm the high purity and integrity of the peptide sample. These results support the use of PS77 in further biological studies and its potential development as a therapeutic agent.
Fig. 1
A The predicted 3D structure of PS77 with Van der walls forces. B HPLC spectrum and the corresponding peak intensities of PS77. It showed the chromatographic separation of the peptide sample, with the x-axis representing the retention time and the y-axis indicating the intensity or absorbance of the detected compounds. C MS spectrum and the corresponding peak intensities of PS77. The MS spectrum displayed the mass-to-charge ratio (m/z) of the peptide fragments on the x-axis and their relative abundance on the y-axis. The single and intense peak corresponds to the molecular weight of the PS77 peptide
Furthermore, the α-helical structure of PS77 is pivotal in biomaterial design, conferring enhanced specificity in targeting inflammatory pathways, which is crucial for developing targeted therapies with minimal off-target effects. This structural configuration also contributes to the peptide’s stability and biocompatibility, essential attributes for any biomaterial intended for therapeutic use, ensuring that it remains active and well-tolerated within the biological environment. Moreover, these properties make PS77 an ideal candidate for integration into sophisticated delivery systems or scaffolds for tissue engineering, facilitating localized therapy at inflammation sites while minimizing systemic exposure and potential side effects.
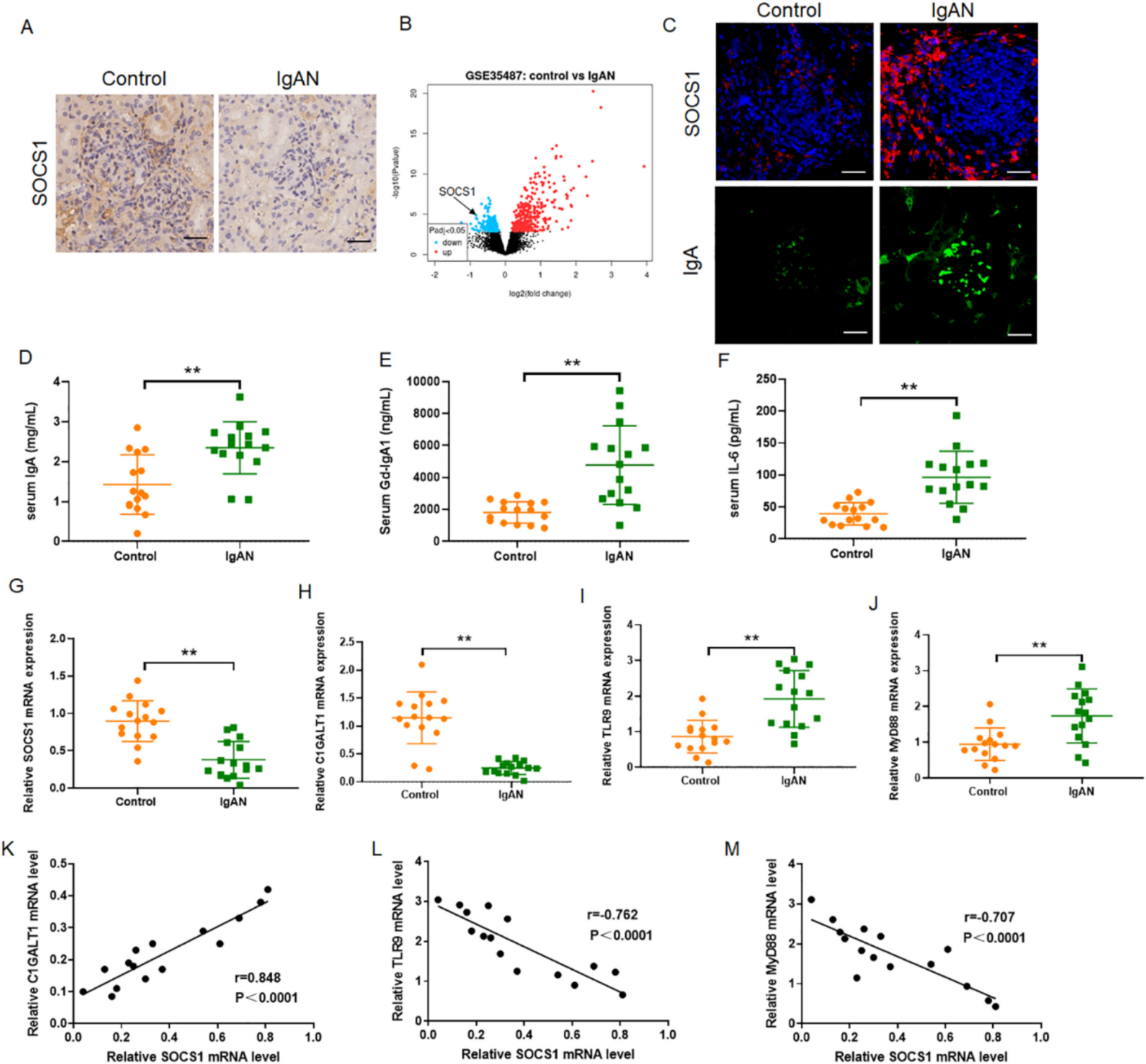
Establishing TNF-α-induced inflammatory modelTo explore the effects of PS77 on inflammation, an in vitro model was established using human keratinocyte (HaCaT) cells treated with tumor necrosis factor-α (TNF-α). Cell viability assays showed no significant differences (P > 0.05) between normal HaCaT cells and those treated with TNF-α (20–140 ng/mL) for 2 h (Fig. 2A). TNF-α at 20 ng/mL was chosen for subsequent experiments. Enzyme-linked immunosorbent assay (ELISA) confirmed significantly increased (P < 0.05) TNF-α levels in the inflammatory model compared to normal controls (Fig. 2B), validating the model’s establishment.
Fig. 2
Effect and cytokine expression in the TNF-α-induced and PS77-treated on HaCaT cells. A Effect of TNF-α across different concentrations on normal HaCaT cells. B Cytokine expression in the TNF-α-induced inflammatory in model. C Effect of PS77 at different concentrations on normal HaCaT cells. D Cytokine expression in PS77-treated TNF-α-induced inflammatory model. The cell viability tests were averaged based on the cell viability of three independent biological replicates and statistically analyzed by Prism software, bar is shown as Mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.0005, ****P < 0.0001. TNF-α: TNF-α-induced inflammatory model, PS77: PS77-treated inflammatory model
PS77 exhibits no significant toxicity on normal HaCaT cellsTo explore the toxicity of PS77, cell viability assay was performed using normal HaCaT cells treated with the PS77 peptide (0.1–100 μg/mL) for 24 h. Our data showed that PS77-treated HaCaT cells in a concentration range of 0.1 to 100 μg/mL did not significantly affect the viability of normal HaCaT cells (cell viability was approximately 100%), suggesting its safety for potential therapeutic use (Fig. 2C). A concentration of 0.1 μg/mL was selected for further studies.
To examine the anti-inflammatory activity of PS77, we treated inflammatory model with 0.1 μg/mL of PS77 and examined the expression of the sets of inflammatory markers using ELISA assay. Our analysis revealed that PS77 treatment (0.1 μg/mL) significantly reduced (P < 0.05) the expression of pro-inflammatory cytokines IL-8 and MMP-3 by 20–30% in the TNF-α-induced inflammatory model, indicating PS77's anti-inflammatory effects (Fig. 2D). The pro-inflammatory cytokine expression rates were reduced 20 to 30% after the PS77 treatment, indicating that the anti-inflammatory effect of PS77 might be through the downregulation of the expression of these pro-inflammatory cytokines.
Transcriptomic analysis of PS77-treated inflammatory modelTo elucidate the mechanisms underlying PS77's anti-inflammatory activity, whole-transcriptome sequencing was performed on the PS77-treated inflammatory model (five biological replicates) and untreated controls. High-quality RNA samples (Supplementary Table 2) yielded 722.7 million paired-end reads (148 bp) with > 94% Q30 scores (Supplementary Table 3). Reads were mapped to the human reference genome using Hisat2(v2.0.1)1, with mapping rates > 92%, indicating the high quality of our transcriptomic data and is suitable for downstream analyses (Supplementary Table 4).
Gene expression analysis revealed similar profiles between PS77-treated and control samples (Fig. 3A–B), suggesting few differentially expressed genes (DEGs) upon treatment. However, 265 significant DEGs (FC ≥ 2, P < 0.05) were identified, with 137 upregulated and 128 downregulated genes (Fig. 3C–D, Table 1 & Supplementary Table 5). Several of the downregulated genes are related to inflammatory and immune processes (Table 1). For instance, the key inflammation-related gene CHRNA7 is a cholinergic receptor that can influence inflammatory responses [27]. INHBB and CXCR5, cytokine and chemokine receptors involved in immune cell signaling and migration [28], were also downregulated. CAND2 and KLK4 are involved in cullin-associated protein degradation, which can modulate inflammatory signaling pathways. Notably, genes involved in the bone morphogenetic protein (BMP) signaling pathway, such as BMP5 and DLX5, were downregulated. The downregulation of these genes by PS77 aligns with its anti-inflammatory effects, as reducing the expression of pro-inflammatory genes would help dampen inflammation. Some upregulated genes also have potential anti-inflammatory roles. RXRG is a retinoid X receptor gamma, which can form complexes that inhibit inflammatory gene expression [29], whereas both IL12RB2 [30] and IL2RG are subunits of interleukin receptors that modulate interleukin signaling and can have immunoregulatory effects [31, 32], while anti-inflammatory genes.
Fig. 3
A FPKM density distribution correlation plots. B Box plots of gene expression level for each sample. C Volcano plot of DEGs regulated by PS77. D Heatmap of DEGs in different samples
Table 1 List of selected DEGsFunctional enrichment analysis of DEGsTo have better insights into the functions of these downregulated DEGs, we performed the functional enrichment analysis using the provided tool on the Gene Ontology website (http://geneontology.org/). Our data revealed that downregulated DEGs were involved in biological processes such as response to BMP and cellular response to BMP stimulus, with genes including DLX5, BMP5, ELAPOR2, and ADAMTS7 (Fig. 4A). BMPs are members of the TGF-β superfamily and have been implicated in various inflammatory processes [33]. DLX5 has been reported to regulate chondrocyte hypertrophy [34], which is associated with osteoarthritis, a chronic inflammatory condition. By downregulating DLX5, PS77 may help to mitigate inflammation in osteoarthritis. However, the downregulation of genes involved in BMP signaling, such as DLX5 and BMP5, suggests that PS77 may exert its anti-inflammatory effects by modulating the BMP signaling pathway. Molecular functions enriched among downregulated DEGs included BMP receptor binding, transmembrane receptor protein serine/threonine kinase binding, receptor serine/threonine kinase binding, and structural constituent of muscle, with genes such as BMP5, ELAPOR2, MYH11, and SORBS2 (Fig. 4A).
Fig. 4
A GO biological process and molecular function enrichment analysis of all the downregulated genes. B KEGG pathway analysis of downregulated DEGs regulated by PS77
Pathway enrichment analysisTo understand and demonstrate the biochemical metabolic pathways and signal transduction pathways involved in DEGs, we performed pathway analysis using the provide tool on the KEGG [35]. Our analysis showed that the anti-inflammatory peptide PS77 downregulated genes that were enriched in several key pathways related to inflammation and cell signaling. The most significantly enriched pathway was the cytokine-cytokine receptor interaction, with a gene ratio of approximately 0.25. Cytokines are crucial mediators of inflammation, and their interaction with specific receptors triggers various signaling cascades that regulate the inflammatory response. The downregulation of genes involved in this pathway by PS77 suggests that the peptide may modulate the signaling of various cytokines, which are known to play crucial roles in inflammation [36]. By reducing the expression of these pro-inflammatory cytokines and their receptors, PS77 may help to dampen the inflammatory response. Another important pathway downregulated by PS77 was the TGF-beta signaling pathway, with a gene ratio of around 0.15 and involving genes such as BMP5, INHBB, and CXCR5 (Fig. 4B). The TGF-beta superfamily, which includes bone morphogenetic proteins (BMPs), plays a significant role in regulating inflammation, cell proliferation, and differentiation. INHBB, a member of the TGF-β superfamily, has been shown to promote the activation of fibroblasts and the production of pro-inflammatory cytokines [37]. By downregulating INHBB, PS77 may help to reduce inflammation and fibrosis. PS77 also downregulated genes involved in the 2-oxocarboxylic acid metabolism, arginine biosynthesis, and vitamin digestion and absorption pathways, although to a lesser extent (gene ratios < 0.10). For instance, arginine metabolism has been linked to the regulation of immune responses and inflammation [38]. By altering arginine biosynthesis, PS77 may influence the availability of this amino acid for immune cell function and cytokine production. These pathways are involved in various cellular processes, and their modulation by PS77 may indirectly contribute to its anti-inflammatory effects by altering cellular metabolism and nutrient availability.
Altogether our functional and pathway enrichment analyses provide valuable insights into the potential mechanisms underlying the anti-inflammatory effects of PS77. By modulating key inflammatory pathways, such as BMP and TGF-β signaling, and by regulating the expression of pro-inflammatory cytokines and their receptors, PS77 may exert a potent anti-inflammatory action. These findings support the further development of PS77 as a novel therapeutic agent for inflammatory diseases.
Comments (0)