
Remember me
TUT4 and TUT7 are multidomain enzymes whose specific targeting of RNAs makes use of three zinc fingers, combined with two copies of a nucleotidyltransferase domain (NTD), one inactive in the N-terminal half of the polypeptide (pseudo-NTD) and one active in the C-terminal half (Fig. 1a,b)4,5. Structures have previously been determined for the N-terminal LIN28A-interacting module (LIM) of TUT4 including its pseudo-NTD domain33, and for the C-terminal catalytic module (CM) of TUT7 (ref. 34), as well as for LIN28A bound with RNA constructs representing pre-let-7d/f/g miRNAs35,36. The human TUT4 LIM33 was found to have a very similar basic architecture to that of the TUT7 CM34. The TUT4 LIM structure was, however, distinguished by the presence of its zinc finger domain at the N terminus, which is homologous to those of double-stranded RNA-binding zinc fingers33. The TUT7 CM structure was previously solved crystallographically in apo and complex forms with double-stranded RNA, U2, U5 or uridine triphosphate (UTP)34. In these CM structures, the second TUT zinc knuckle (ZK2) could be visualized bound with both U2 and U5. The diverse substrate complexes revealed that the RNA-binding pocket orients a group II pre-let-7 to favor mono-uridylation, while the ZK2 of TUT7 is thought to aid oligo-uridylation by supporting the extension of the oligo-U tail34. However, none of the structures determined so far have been able to show the mechanistic basis for the alternative activities shown by TUT7 and TUT4: that is, mono- versus oligo-uridylation.
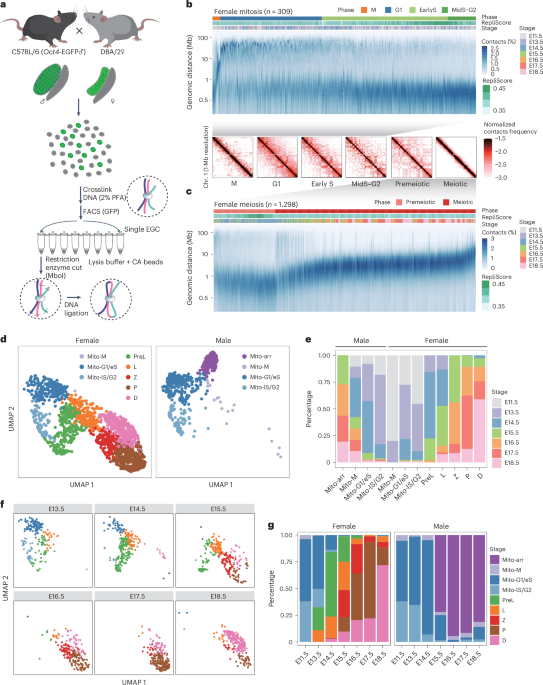
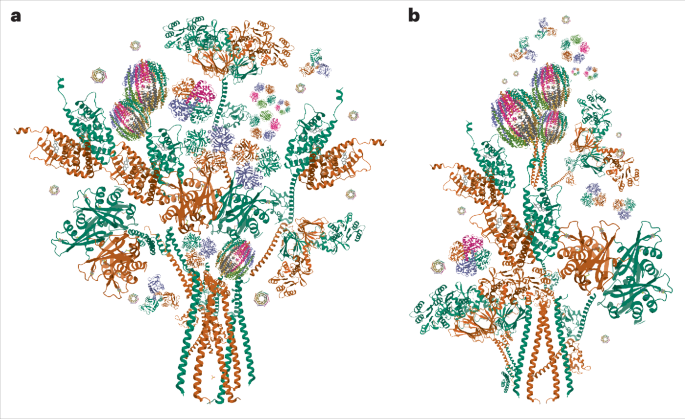
Fig. 1: Cryo-EM structures of TUT7 in apo and binary complex with pre-let-7.
a, Human TUT7 domain organization. b, Human TUT4 domain organization. c, Human LIN28A domain organization. d, Nucleotide sequence of pre-let-7g, presented in a predicted secondary structure. e, Overall structure of TUT7 alone, shown as a 4.03 Å cryo-EM density map with cartoon representation of protein backbone, in two views related by a 90° rotation, as indicated. A handed model of TUT7 apo is presented at the upper right corner. The canonical palm and finger domains of the NTD canonical fold are colored as follows, N-terminal pseudo-NTD (from LIM) in red and pink and C-terminal NTD (CM) in blue and cyan. f, Overall structure of the TUT7/pre-let-7g binary complex, shown as a 3.55 Å cryo-EM density map with cartoon representation of the fitted coordinates in two views related by a 150° rotation, as indicated. TUT7 colored as in e, ZK1 colored orange, pre-let-7g yellow. g, Cartoon model of TUT7 in complex with pre-let-7g, highlighting the relative arrangements of domains in a C-shape with the pre-miRNA bound. h, Zoomed-in view of UTPαS within the catalytic pocket, interacting with the terminal pre-let-7g base C78 and the TUT7 residues shown. i, Close-up of RNA binding by ZK1, highlighting the role of the K969 side chain interacting with G5 and U6 within the pre-let-7g stem.
LIN28, as a key regulator controlling the timing of developmental events and participating in heterochronic processes, is highly conserved across diverse organisms37. In C. elegans, there is only one encoded LIN28 protein, while in humans there are two paralogs, LIN28A and LIN28B17,37. LIN28 proteins are composed of a cold-shock domain (CSD) and a zinc knuckle (ZK) domain belonging to the zinc finger CCHC-type (ZCCHC) superfamily (Fig. 1c).
To better understand the uridylation mechanism involved in the biogenesis of mature let-7, we used single-particle cryo-EM to study the structures of TUT7 and TUT4 without and with pre-let-7g miRNA (Fig. 1d, see Supplementary Information for RNA sequences), in the absence or presence of LIN28A. We purified recombinant full-length wild-type human TUT7 and human LIN28A from Escherichia coli strain KRX38. Synthetic pre-let-7g was used as a substrate for structural studies, because it has been widely used in previous studies of TUT4/7 activity4,34. To visualize the TUT7 apo state we initially isolated the enzyme bound with UTPαS alone (Extended Data Fig. 1a,b) but found that modification with NHS-polyethyleneglycol4 (NHS-PEG4)39 was needed to avert excessive sticking of particles to the air–water interface where they are prone to denaturation (Extended Data Fig. 1b,c). Unmodified enzyme was combined with pre-let-7g to obtain the binary TUT7/pre-miRNA complex (Extended Data Fig. 1b,d) and the TUT7/LIN28A/pre-let-7g ternary complex (Extended Data Fig. 1b,e). The activity of TUT7 as expressed here and assembled with pre-let-7g miRNA was confirmed both without and with LIN28A, resulting in mono- and oligo-uridylation, respectively (Extended Data Fig. 1f). Using the isolated apo form and assembled sample complexes we embedded the TUT7 samples in vitreous ice and solved a series of cryo-EM structures in different functionally related states and with overall resolutions ranging from 3.53 to 4.03 Å (Table 1). To compare with TUT7, a truncated TUT4 (residues 254–1315) with a similar activity to the full-length TUT4 protein and to an N-terminal-only truncated form (residues 153–1649) was expressed and purified from the E. coli KRX strain and assembled with LIN28A and pre-let-7g before polishing by size-exclusion chromatography (Extended Data Fig. 1g–i). As before, enzyme activity for the truncated TUT4 construct was confirmed both without and with LIN28A (Extended Data Fig. 1j). We thereby succeeded in obtaining a cryo-EM map of the TUT4 ternary complex at a resolution of 3.68 Å (Table 1).
Table 1 Cryo-EM data collection, refinement and validation statisticsLoose association between NTDs characterizes the TUT7 apo stateThe TUT7 structure in an apo state at 4.03 Å revealed an overall ‘C-shape’ conformation composed of two right-handed conformers (Fig. 1e and Extended Data Fig. 2a–f) distinct from the ‘L-shape’ of human Dicer, which in addition remains similar in its conformation throughout its activity cycle14. This difference suits Dicer’s role as a ‘tape-measure’ protein capable of recognizing the length associated with a correctly generated pre-miRNA, hence its need for a fixed dimension, whereas the TUTs need to be able to function in two different modes (mono- and oligo-uridylation) and, further, to be able to act processively on a bound pre-miRNA substrate under the influence of LIN28A.
TUT7 alone binds pre-let-7g miRNA via end-capture of the target 3′ endA 3.55 Å map of TUT7 in a binary complex with a pre-let-7g (Fig. 1f,g and Extended Data Figs. 2g–l and 3a) shows the RNA stem bound into the catalytic cleft of its CM, with the cytosine single-nucleotide overhang at the pre-miRNA 3′ end (C78) and UTPαS bound and stabilized by the critical residues; F1045, S1047, Q1124, L1126, N1130, K1156, S1170, Y1171 and H1286 (Fig. 1h and Extended Data Fig. 3b) (H1286 is equivalent to H336 in the yeast homolog that is known to be critical for uracil selection40,41,42). The let-7g pre-element is oriented away from the CM and, lacking contact with ordered protein structure, was found to be relatively disordered, presumably due to some intrinsic flexibility. Interpretation of contacts made by TUT7 with the RNA stem was assisted by modeling of its ZK1, which fitted well into a clearly visualized but relatively low-resolution region of density (Fig. 1f and Extended Data Fig. 3c,d). The ZK1 region had not been resolved in the previous structural study of the CM alone34, which instead provided an atomic model for ZK2, a region of TUT7 that supports oligo-uridylation by engaging the extending U tail34, but is not represented in our structure due to disorder.
In both the apo state (Fig. 1e) and the binary TUT7 complex (Fig. 1f,g) the two NTDs maintain the distinctive ‘C-shape’, apparently due to their direct interaction with one another. The interface between the LIM and CM is occupied by several nonpolar and polar amino acids, indicating the likelihood of a hydrophobic lateral interaction, as observed also in Dicer through the RNase IIIa and IIIb domains14. The binding of the pre-miRNA results in a higher-resolution structure with more of TUT7 properly resolved in the cryo-EM density, and with an extensively extended polypeptide region linking the LIM and CM that includes its ZK1 region (Extended Data Fig. 3d). Thus partially stabilized, the binary TUT7/pre-miRNA complex shows a defined but dynamic interface between the LIM and CM domains (Supplementary Video 1). By contrast, in the apo state no obvious density was resolved for the backbone link connecting the LIM and CM, with only a small region of the linking density visible associated with the LIM (Extended Data Fig. 3e). Nevertheless, for maintenance of the distinctive ‘C-shape’ structure, the two NTDs of the enzyme must still be interacting directly in the apo state, albeit relatively weakly. These two models show us the conformation of TUT7 (and we posit, TUT4) in the state before substrate binding (Fig. 1e) and when a pre-miRNA alone associates with it (Fig. 1f,g).
The roles of all three TUT4/7 ZKs were probed in a recent NMR-based study that made use of pre-let-7i as a target43. While binding of TUT4 ZK2 and ZK3 to RNA was shown, it was concluded that ZK1 had lost its ability to interact with RNA due to two key mutations within its CCHC motif after the first cysteine and the single histidine, where the relevant amino acids were mutated from a variable bulky mostly hydrophobic residue to a serine43. Alignment of the amino acids of the three ZKs of TUT7 (Extended Data Fig. 3f) showed the residue after the first cysteine of ZK1 was serine, but the one after its histidine was leucine, different from the situation in TUT4 where it is another serine43. Nevertheless, in the binary complex we resolve, the lysine (K969) after the second TUT7 cysteine of ZK1 is shown to interact with the G5 and U6 of pre-let-7 using its side chain (Fig. 1i and Extended Data Fig. 3c). This reveals that one of the functions for ZK1, at least in TUT7, is to recruit or stabilize pre-let-7 for mono-uridylation through a direct interaction with its linker loop. K969 of TUT7 is conserved in the ZK1 of TUT4 as K919 (Extended Data Fig. 3f).
LIN28A modifies target pre-miRNA recognition by previous associationTwo distinct conformations (I and II) of the TUT7 ternary complex were resolved at 3.81 and 3.53 Å resolutions, respectively (see Extended Data Fig. 4 for the data processing pipeline and Extended Data Fig. 5 for close-ups of local density). In conformation I (Extended Data Figs. 4a–f and 5a), most of the pre-let-7g could be resolved (Fig. 2a,b). In this conformation, the RNA stem interacts with the TUT7 LIM, while the let-7g 49AGGAG53 region coordinates with the zinc finger of TUT7 and ZK domain of LIN28A, forming a stable ternary complex35 (Fig. 2a,b). Furthermore, despite clear density representing the pre-element (the apical loop and short stem region preceding later removed by Dicer action), no density was observed for the CSD of LIN28A, presumably due to the relative flexibility between the LIM + LIN28A and pre-let-7g. Based on the position of the RNA, this complex is believed to represent the initial step in oligo-uridylation, a ‘capture’ state in which the pre-let-7 has begun association with TUT7 but not yet completed its engagement.
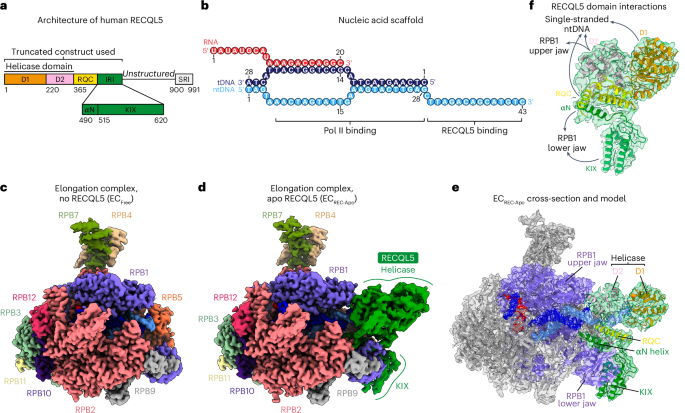
Fig. 2: Cryo-EM structures of TUT7 in complex with pre-let-7g and LIN28A.
a, Overall structure of the TUT7/pre-let-7g/LIN28A ternary complex conformation I, shown as a 3.81 Å cryo-EM density map and cartoon representations. C-terminal NTD (CM) fingers in cyan and palm in blue; the LIM zinc finger is colored sand, LIN28A ZK dark orange and pre-let-7g miRNA yellow, in two views related by a 90° rotation. b, Cartoon model of TUT7/pre-let-7g/LIN28A ternary complex conformation I, colors as in a. c, Overall structure of the TUT7/pre-let-7g/LIN28A ternary complex conformation II, shown as a 3.53 Å cryo-EM density map and cartoon representation. Colors as in a and b, plus TUT7 ZK1 and linker region in bright orange, and pseudo-NTD from the LIM (fingers red and palm pink), in two views related by a 90° rotation as indicated. d, Cartoon model of TUT7/pre-let-7/LIN28A ternary conformation II. Areas shown in expanded views below are boxed. e, Zoomed-in, two views of domain interactionn between TUT7 LIM and TUT7 CM related by a 120° rotation as indicated. f, Close-up of TUT7 ZK1 bound to the pre-let-7, highlighting key roles for K969 and for K970, E971 and E980, among other residues in two views related by an 80° rotation.
In conformation II (Fig. 2c,d and Extended Data Figs. 4a–c,g–i and 5b), the CM density missing in conformation I due to intrinsic flexibility could be entirely resolved as well as the LIM-CM linker region including the ZK1 domain (Extended Data Fig. 6a). The higher-resolution model enabled us to observe intramolecular interactions at a residue level, especially for the interface between the LIM and CM. For example, a hydrophobic interaction formed by ring-stacked residues F389 and F1267, and a H390-induced polar interaction with T1266 and T1272 stabilize the interface (Fig. 2e and Extended Data Fig. 6b), thus also confirming the role of a similar interface in the TUT7 binary complex (Fig. 1f), and presumably the apo state also. Together, these interactions ensure maintenance of the distinctive conformation of TUT4/7. In this state of the ternary complex R970 forms two hydrogen bonds with U13 and U14 in the RNA stem while K969 binds the sugar backbone of G66 (Fig. 2f and Extended Data Fig. 6c), which is different from the binary complex in which its side chain instead interacts with G5 and U6 (Extended Data Fig. 3c). In addition, residues E971 and E980 from ZK1 further stabilize RNA binding by interactions with the pre-let-7g backbone (G15) and the base of U13 (Fig. 2f). Altogether, these interactions aid the translocation of pre-let-7 to close the catalytic pocket of the CM and stabilize the TUT7 bound to the pre-miRNA and LIN28A.
Both models of ternary complexes revealed the ZK domain of LIN28A attached to its 49AGGAG53 recognition sequence in the pre-let-7 together with the zinc finger of the TUT7 LIM. To explore the potential conformational changes in the overall structure, a three-dimensional (3D) conformational variability analysis was performed (Supplementary Video 2). The TUT7 LIM and LIN28A were found to be stable, while the pre-element of pre-let-7g miRNA exhibited density that could be assigned to a set of flexibly related conformations. The angle between the pre-element and RNA stem changed from around the initial (frame no. 1) 70° to a final (frame no. 20) 40°. The finding was repeated via the same analysis for the ternary complex in conformation II (Supplementary Video 3). This result is consistent with a previous study, in which it was concluded that in solution, the pre-let-7 alone may adopt different conformations with various stem stabilities14, which we now observe in our cryo-EM structures bound to TUT7. Since the CSD was thought to bind in the pre-element region this may explain why this density could not be resolved.
Altogether, comparison of the two TUT7 ternary complexes and the binary and apo structures (Fig. 3a–g) indicates that the presence of LIN28A has changed how the pre-let-7 engages with the enzyme, directing it away from direct binding to the CM active site (as in the binary complex) and toward a two-stage process in which, first, the LIM and LIN28A capture the stem and pre-element, and then the CM engages with the 3′ end. Comparison of the apo TUT7 and its ternary complex conformation II indicates a 7.5° rotation of the LIM region with respect to the CM, while the switch from binary TUT7 to ternary conformation II involves less than half of this movement (2.7°); additionally, the Nc-palm-1 moves 4.4° with respect to the Fingers-1 subdomain, within the LIM, as the TUT7 binary complex converts ultimately to ternary conformation II (Fig. 3d,e). By contrast, comparison of TUT7 ternary conformations I and II finds no change in the relative positions of the palm and fingers subdomains of the LIM while the zinc finger binding the pre-let-7 stem moves substantially about a 16.8° rotation of the linking helix (Fig. 3f,g). Further details of the first of these steps—formation of the capture complex—were provided by determination of the equivalent structure for TUT4, also with LIN28A and pre-let-7g.
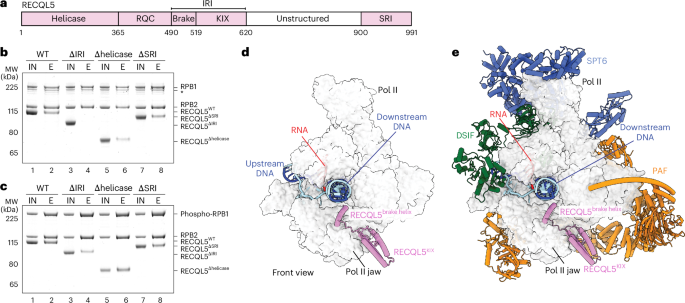
Fig. 3: Structural comparison of TUT7 cryo-EM structures.
a–c, Cartoon models of TUT7 in apo state (a), as a binary complex with pre-let-7g (b) and in ternary complex with pre-let-7g and LIN28A (conformation II) (c). Domains are colored as in the main figures and zinc ions are blue spheres. d, Superimposition of the TUT7 apo (white) and pre-let-7g bound cryo-EM structure from the TUT7/pre-let-7g/LIN28A ternary complex conformation II (colored), aligning by the TUT7 CM. The reorientation of the LIM results in a ~5 Å shift toward the pre-miRNA, as shown. Angular changes in this figure were calculated by reference to the centers of mass of the respective regions, marked here by red dots. e, Superimposition of the TUT7/pre-let-7g binary complex (deep gray) and the TUT7/pre-let-7g/LIN28A complex conformation II (colored), in two views related by 60° and 90° as shown, and aligning by the TUT7 CM. f, Cartoon model of TUT7/pre-let-7g/LIN28A ternary complex conformation I. g, Superimposition of the TUT7/pre-let-7g/LIN28A complex conformation I (pale green) and conformation II (colored), aligning by the TUT7 LIM.
TUT4 capture complex determined at higher resolutionA 3.68 Å cryo-EM structure was obtained for the TUT4/LIN28A/pre-let-7 in the pre-oligo-uridylation capture state (Extended Data Fig. 7) found also as TUT7 ternary conformation I. This structure enabled us to identify more clearly the interactions between the LIM module of TUT4, LIN28A and the pre-let-7 (Fig. 4a). Although the CM of TUT4 is not well defined in the 3D map, density apparently derived from it can be visualized in two-dimensional (2D) class averages of the complex, roughly oriented in the same view compared to the TUT4 ternary complex in conformation I (Fig. 4b), in line with this being a capture state before CM engagement with the open end of the pre-let-7g. The 49AGGAG53 recognition sequence from pre-let-7g was observed trapped into a positively charged pocket created by TUT4 and LIN28A (Fig. 4c,d). Together with the TUT7 ternary complexes this structure therefore provides a demonstration that TUT4 interacts directly with LIN28A, a currently unresolved and key question in the field. In this ternary interface, a hydrophobic patch is formed between G276, L277, A280, I316 from TUT4 LIM (which are conserved in TUT7: G216, L217, A220, I256) and prolines 156 and 168 of LIN28A (Fig. 4e,f). Overall, the structures of the TUT4 and TUT7 LIMs are very similar (root mean squared deviation 1.22 Å over 302 core backbone Cα atoms, 2.18 Å over all 380 pairs) (Extended Data Fig. 8a,b). Compared to the crystal structure of TUT4 LIM33 (Extended Data Fig. 8c), the angle between the helices α4 in the zinc finger and α5 in the palm subdomain is reduced by 12.6° during the binding of pre-let-7g (Extended Data Fig. 8d).
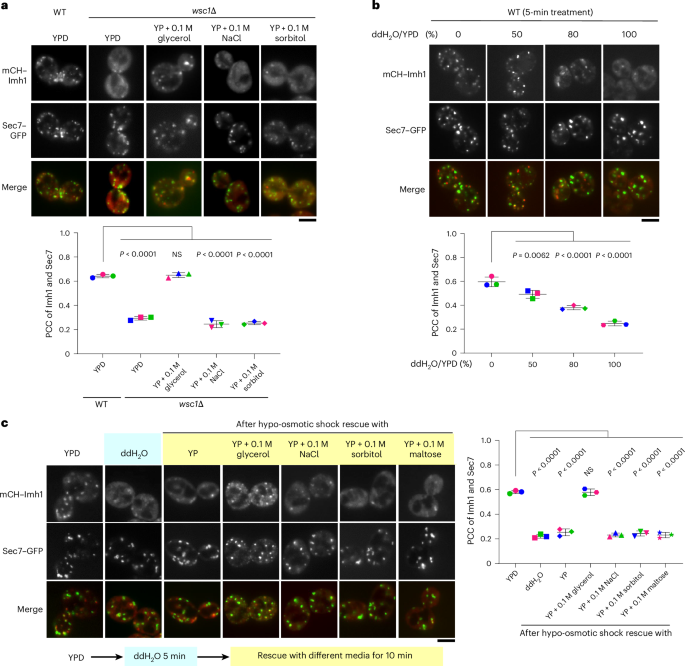
Fig. 4: Cryo-EM structure of TUT4 in complex with pre-let-7g with LIN28A.
a, Overall structure of TUT4/pre-let-7g/LIN28A, shown as a 3.68 Å cryo-EM density map and cartoon representations in three views, related by 90° and 180° rotations, as shown. Colors are as previously: pseudo-NTD (LIM) fingers and palm in cyan and blue, respectively; LIM zinc finger and linking density toward CM is colored sand, pre-let-7g yellow and LIN28A ZK dark orange. b, A typical 2D class average representing the poorly resolved density from the CM (marked by a red arrowhead). c, Electrostatic surface potential of TUT4/pre-let-7g/LIN28A. The positively and negatively charged regions are colored from blue to red, respectively. d, Zoomed-in view of the pre-let-7g 49AGGAG53 region in a positively charged pocket created by the TUT4 zinc finger and ZK domain of LIN28A (black arrows). e, Cartoon model of TUT4/pre-let-7g/LIN28A ternary complex, colored as in other panels. f, Zoomed-in view of TUT4/LIN28A interface. Gray dashed lines link atoms within 3.8 Å and the pale pink dashed line shows a distance (at G276) between two atoms of 3.8–4 Å.
The pre-let-7g is observed bound to three main interfaces in the TUT4 capture complex (Fig. 5a), which we have termed interface 1, interface 2 and interface 3, respectively (Fig. 5b–f). In interface 1, the TUT4 LIM P667, R669, R670 and N688 interact with A64 and G65 at their deoxyribose and phosphate group (Fig. 5c). The pre-let-7 A17 and C18 backbone regions are bound by R327 and K321, respectively, in the second interface region, while the U16 backbone interacts with L691 (Fig. 5d). Interface 3 is critical to the ternary interaction: in this region the zinc finger of the TUT4 LIM is observed to bind the 49AGGAG53 region of pre-let-7 and the two ZKs of LIN28A. The side chains of four positively charged residues R283, R286, H320 and K324 from the LIM participate in the interaction with G51, A52, G53 and A54 of the pre-let-7g (Fig. 5e,f).
Fig. 5: Pre-let-7 recognition by TUT4 and LIN28A.
a, A schematic of pre-let-7 recognition by TUT4 and LIN28A, based on the structure reported here (also Fig. 4). b, Structure of TUT4/LIN28A bound to pre-let-7g. c–f, Zoomed-in views of the different interfaces bound to pre-let-7g: interface 1 (c), interface 2 (d) and interface 3 (e and f). Throughout this figure, gray dashed lines link atoms within 3.8 Å and pale pink dashed lines show a distance between two atoms of 3.8–4 Å.
The two ZKs of LIN28A interact with the 49AGGAG53 motif of pre-let-7g, in a similar way to the previously solved isolated complexes35,44. Based on these structures, it was concluded that the double ZK motif recognizes two AG dinucleotides separated by a single-nucleotide spacer35,44. Perhaps due to the use of a modified RNA sequence mimicking the full pre-let-7g, the exact basis for recognition observed differed from the one we identify. In the previously solved crystal structures the LIN28A ZK1 binds with the A54 of pre-let-7g via a hydrogen bond from the Y140 sidechain, whereas we observe the tyrosine binding with G51, A52 and G53 (Fig. 5a,e). In all, the pre-let-7 miRNA is sandwiched between LIN28A and the TUT4 LIM (Figs. 4c,d and 5b–f and Extended Data Fig. 8e) as it was in the TUT7/pre-let-7g/LIN28A complex (Extended Data Fig. 8f,g) and so as to engage with the pre-miRNA in a fundamentally different way from that observed for the binary TUT7/pre-let-7g complex set for mono-uridylation (Extended Data Fig. 8h). The binding mode and overall conformation of the pre-let-7g miRNA appears to be remarkably consistent in all three LIN28A-containing complexes (Extended Data Fig. 8e–g), apparently due to the LIM/LIN28A clasp (Fig. 5f) and other key interfaces shown by mutational data to be the specific interactions responsible for ensuring efficient and processive oligo-uridylation.
Mutagenesis of TUT4 (Fig. 6a) in interfaces 2 and 3 still enabled addition of a single uridine to pre-let-7g in the absence of LIN28A (Fig. 6b), but abolished the enzyme’s capacity to oligo-uridylate pre-let-7 in the presence of LIN28A (Fig. 6c–e) due to a much-reduced ability to form a TUT/pre-let-7g/LIN28A ternary assembly (Fig. 6f,g). Although mutations in the interface 1 affected the formation of the ternary complex (Fig. 6f,g), they did so less than interface 2 and 3 mutations, and still permitted oligo-uridylation, albeit with a lower efficiency compared to wild type (Fig. 6c–e). No single-site mutations had a notable impact on TUT4 activity, hence the use of double mutants in the assays shown.
Fig. 6: Effects of mutants at the TUT4/pre-let-7g interface on uridylation activity.
a, SDS–PAGE of recombinant TUT4 wild type and its mutants. This expression trial was performed twice. b, In vitro uridylation of pre-let-7g by wild-type TUT4 and mutants, in the absence of LIN28A, as indicated. Pre-let-7, substrate, indicated as Marker1, 78 nt, shown in lane 2 (see Supplementary Information for RNA sequence, pre-let-7g); pre-let-7-U, synthetic substrate, indicated as Marker2, 79 nt, shown in lane 1 (see Supplementary Information for RNA sequence pre-let-7g_1U); Mono-U pre-let-7g, pre-let-7g products with a single U addition; Oligo-U pre-let-7g, pre-let-7g products with more than two U addition. K919 and K920 mutated here are the equivalent residues to K969 and R970 in TUT7 (Fig. 2f). This assay was performed three times. c, Relative oligo-uridylation activity of wild type or the indicated mutant TUT4 enzymes in the presence of LIN28A. Markers as for b. This assay was performed three times. d, In vitro uridylation of pre-let-7g by TUT4 and its variants in the presence of LIN28A. Error bars show the standard error of mean, n = 3 independent experiments (trials 1, 2 and 3) (see Source data for the raw data plotted). e, Time course activity assay for wild-type and mutant forms of TUT4 in the presence of LIN28A; Marker1, pre-let-7, 78 nt, shown in lane 1. Two repeats were performed for this assay. f, EMSA of pre-let-7g with wild-type and mutant forms of TUT4 only. This assay was performed twice. g, EMSA of pre-let-7g with wild-type and mutant forms of TUT4, combined with LIN28A and TUT4. This assay was performed twice. h, EMSA of pre-let-7g with increasing concentrations of LIN28A alone, TUT4 alone and LIN28A + TUT4 together. Marker1, pre-let-7, 78 nt, 25 kDa, shown in lane 1. This assay was performed twice.
To our surprise, mutations in the ZK1 region that binds to the RNA stem in the TUT7 ternary conformation II, had no effect on oligo-uridylation activity (Fig. 6c–e), indicating other residues play critical roles in RNA binding, for example D921 and E930 that are the equivalent residues to TUT7 E971 and E980 (Fig. 2f and Extended Data Fig. 9). It is notable that the key interactions we observed in TUT4 complexes involve residues conserved in TUT7 (for example, K919/K920 in TUT4 equivalent to K969/R970 in TUT7), with the one notable difference that L691 in human TUT4 is P613 in TUT7, and is a polypeptide locus in interface 1 besides U16 of pre-let-7g; and otherwise only the conservative substitution of R669 in human TUT4, for K591 in TUT7 (Extended Data Fig. 9). Higher-order assemblies of TUT4/pre-let-7g/LIN28A can also be observed in the shift assays testing complex assembly, for wild-type TUT4 and with mutations at interface 1 and ZK1 (Fig. 6f,g), and we confirmed a concentration-dependent formation of these larger complexes by titrating pre-let-7g with LIN28A and TUT4 alone and together (Fig. 6h).
Comments (0)