
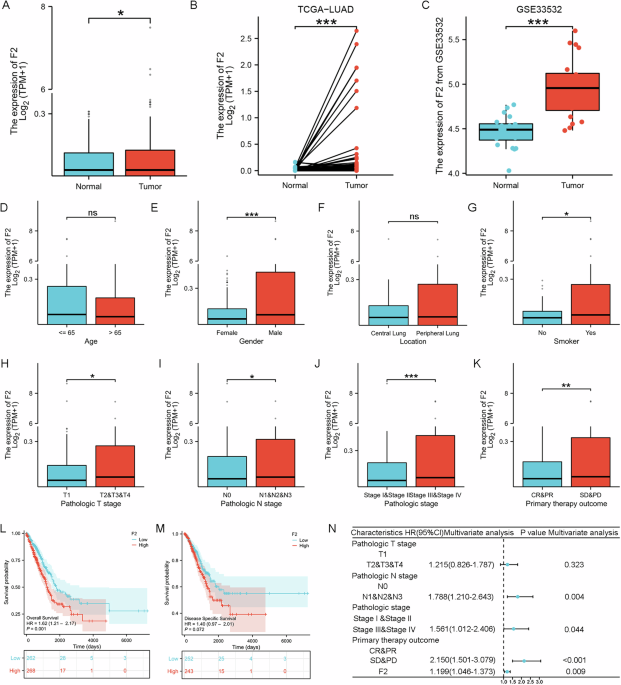
Remember me
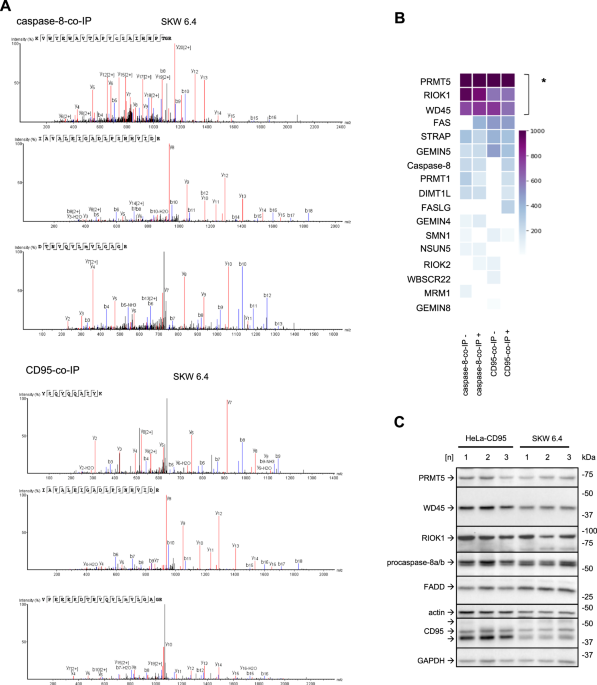
Mass spectrometry analysis of CD95-co-immunoprecipitation (CD95-co-IP) and caspase-8-co-IP from CD95L-stimulated B lymphoblastoid SKW 6.4 cells has identified the presence of the core components of the PRMT5 methylosome complex: PRMT5, WD45 and RIOK1 in these co-IPs (Fig. 1A, B; Supplementary Fig. 1A–C). These three proteins were detected with high confidence in both the CD95-co-IP and the caspase-8-co-IP (Fig. 1A, B; Supplementary Fig. 1A–C). Caspase-8 and CD95/Fas were found in CD95-co-IP and caspase-8-co-IPs, respectively, only upon CD95L stimulation, indicating the specificity of the DISC formation (Fig. 1B). The other core component of the DISC, FADD, was not detected in this assay. FADD is very difficult to detect in the mass spectrometry analysis of the DISC due to the lower abundance of FADD compared to caspase-8 in this complex, as shown previously [8, 9]. The other proteins from the human methylome besides PRMT5, WD45 and RIOK1 were also found in this mass spectrometry analysis, but with the lower scores (Fig. 1B).
Fig. 1: Mass spectrometry analysis identifies the core components of the PRMT5 complex in caspase-8 complexes by mass spectrometry.
A Mass spectrometry identification of unique proteins in caspase-8 co-Immunoprecipitations (caspase-8-co-IP) and CD95-co-IPs. Mass spectra corresponding to the analysis of PRMT5 in caspase-8-co-IP (upper part) or CD95-co-IP (lower part) after CD95L treatment are shown. Three independent replicates are presented. The SKW 6.4 cells were stimulated for 15 minutes with 166 ng/ml of CD95L. In these experiments, anti-caspase-8 and anti-CD95 (anti-APO-1) antibodies were immobilized at the beads, followed by co-immunoprecipitation (co-IP). The co-IPs from untreated and CD95L-treated SKW 6.4 cells were subjected to mass spectrometry analysis. B Results of mass spectrometry analysis for caspase-8-co-IP and CD95-co-IP experiments. The median of the unique peptides from three independent experiments is shown for each protein for CD95L-stimulated (+) and non-stimulated (–) samples. The scoring indicates the increase in unique peptides identified for each protein. Proteins with statistically significantly higher scores compared to other identified proteins are highlighted (* - p-value < 0.05). The statistical significance was calculated with a one-sample bootstrap method. Only proteins of the human methylome and core DISC components are shown. C Expression of indicated proteins in HeLa-CD95 and SKW 6.4 cells was analysed by Western Blot using the corresponding antibodies. Shown are three different cell passage numbers (1, 2, 3) indicated as [n]. co-IP co-immunoprecipitation; beads-only pulldown, [n], cell passage number.
PRMT5, WD45 and RIOK1 are expressed in different cancer cell lines such as cervical cancer HeLa-CD95 [35] and B lymphoblastoid SKW 6.4 cells (Fig. 1C). The PRMT5 methylosome complexes have been reported to be localized in the nucleus as well as in the cytosol [29]. However, the PRMT5 complex that comprises RIOK1 is known to reside in the cytosol. Hence, we next tested whether PRMT5, WD45 and RIOK1 could be found in the cytosolic fraction after cell fractionation. Fractionation of SKW 6.4 cells showed that PRMT5, WD45 and RIOK1 were detected in both nuclear and cytosolic fractions with and without CD95L stimulation, suggesting a possible involvement of these proteins in cytosolic cell death pathways (Supplementary Fig. 1D).
The results of the mass spectrometry screening were validated by co-IPs followed by Western Blot analysis (Supplementary Fig. 2A, B). In these experiments, SKW 6.4 cells were stimulated with CD95L, followed by co-IP with anti-caspase-8 antibodies. These experiments elucidated the presence of PRMT5, WD45 and RIOK1 in caspase-8-co-IPs in SKW 6.4 cells (Supplementary Fig. 2A, B). PRMT5, WD45 and RIOK1 were also detected in caspase-8-co-IPs without stimulation. This suggests that these proteins may constitutively interact with procaspase-8. PRMT5 has been reported in some studies to bind non-specifically to beads, but this was not observed in these experiments. Indeed, no association of PRMT5 or WD45 and only a very weak signal for RIOK1 were obtained in control pulldowns, performed using the beads-only approach (Supplementary Fig. 2A, B). In addition, the signals corresponding to procaspase-8 were found in WD45-IP and PRMT5-co-IP (Supplementary Fig. 2C, D), further strengthening the evidence that procaspase-8 could be bound to the PRMT5 methylosome.
Taken together, the components of the PRMT5 methylosome complex were detected in caspase-8-co-IPs, which suggests the putative methylation of procaspase-8.
In silico analysis predicted two putative arginine methylation sites on procaspase-8 that are highly conserved and often mutated in human cancersProcaspase-8 is one of the DISC proteins that can be methylated by PRMT5. Indeed, the in silico analysis of putative PRMT5 methylation sites of procaspase-8 predicted with a high confidence two methylation sites in procaspase-8a at RG motifs: R233/G234 and R435/G436. For methylation site prediction, we used the PRmePRed web tool for identification of methylation sites at arginine residues [36].
Importantly, R233 and R435 were found to be highly conserved (Fig. 2A). The analysis of the evolutionary conservation of the procaspase-8 protein sequence revealed that both positions R233 and R435 in the human sequence are highly conserved in mammals confirming their putative functional role (Fig. 2A). Interestingly, in the case of R435, some species from avian and rodent taxonomic groups encode procaspase-8 with the amino acid changes corresponding to the R435Q substitution in the human sequence. In the case of R233, some organisms from the amphibian and bird species encode the R233H substitution. Thus, this analysis suggests the putative functional role of the R233 and R435 residues, which are common to higher vertebrates, and suggests the selection of amino acid mutations at these positions in the current study.
Fig. 2: In silico analysis predicted two putative arginine methylation sites on procaspase-8 that are highly conserved and often mutated in human cancers.
A Analysis of conservation of procaspase-8 R233 and R435 positions. The corresponding procaspase-8 protein regions for the representative higher vertebrate organisms are shown. The species tree and time points of the lineage diversification is presented on the left. The amino acid residues corresponding to human R233 and R435 residues are highlighted. B The prevalence of caspase-8 mutations in cancer was analysed using COSMIC database. All mutations mapped to their position in caspase-8a primary structure are shown in the upper panel. Three most prevalent mutations at R233 and R435 are shown in the various cancers in the lower panel: R233Q missense mutation is shown in black, R233W is shown in dark green and R435* nonsense mutation is shown in yellow. C The location of R233 and R435 in the three-dimensional structure of the C-terminal part of procaspase-8a is shown. The latter is computed using Alphafold [51]. The active site cysteine of caspase-8 is shown in yellow, while R233 and R435 are shown in blue colors. D Putative methylation residues are shown in procaspase-8a primary structure (upper panel). Procaspase-8a-R233H-pcDNA3 (R233H), procaspase-8a-R435Q-pcDNA3 (R435Q) as well as procaspase-8a-WT-pcDNA3 (WT) were transfected into HeLa-CD95-C8-KO cells (low panel). The transfection of pcDNA3 (vector) and non-transfected cells (KO) were used as negative controls. The protein expression was analysed by Western Blot. E HeLa-CD95-C8-KO cells were transfected as in (D). Cytosolic and nuclear fractions were prepared. The analysis of PARP1 and FADD was used as a fractionation control. The fractionation was analysed by Western Blot. One representative fractionation out of three independent experiments is shown.
Furthermore, the analysis of cancer databases showed that the mutations at these two residues are quite common for caspase-8 in several human cancers (Fig. 2B). The R233 and R435 positions are frequently mutated leading to R233Q/R233W missense mutations and R435* nonsense mutation. This further supports the functional importance of these two arginine residues.
Both arginine residues are located in the C-terminal catalytic part of procaspase-8. R233 is located in the p18 large catalytic domain, whereas R435 is located in the p10 small catalytic domain (Fig. 2C, D). According to the available three-dimensional structures of active caspase-8, the two putative methylation sites are not located in close proximity to the active center of caspase-8, though their methylation might influence the conformational stability of the caspase-8 enzyme (Fig. 2C).
Procaspase-8 mutations at the putative methylation sites inhibit CD95L-induced caspase activity and apoptosisProcaspase-8a mutants R233H and R435Q (C8-R233H and C8-R435Q) were generated at predicted putative methylation sites. The selection of arginine substitutions was validated in silico via structural modeling to avoid perturbing the folding of procaspase-8. For this purpose, amino acid substitutions were selected that preserve protein energy as estimated by the Rosetta energy function [37]. In this way, both the R435Q and the R233H mutations were selected by us in the structural analysis in order to preserve protein function but to prevent methylation.
Procaspase-8a (C8-WT), C8-R233H and C8-R435Q were transiently transfected into HeLa-CD95-Crispr/Cas9 caspase-8-/- (HeLa-CD95-C8-KO) cells (Fig. 2D). The concentration of the caspase-8 plasmids used for transfection was selected to achieve procaspase-8 expression slightly below the endogenous levels in parental HeLa-CD95 cells (Supplementary Fig. 3). This was done in order to avoid any effects due to the strong overexpression of this protein in the subsequent experiments.
The introduction of the R233H and R435Q mutations into procaspase-8a did not lead to changes in its expression or its cytosolic localisation (Fig. 2E), but resulted in the inhibitory effects on the CD95L-induced apoptotic signaling cascade. Indeed, CD95L-induced caspase-8- and caspase-3/7 activities, as measured by caspase-8 and -3/7 activity assays, respectively, were inhibited in HeLa-CD95-C8-R233H and HeLa-CD95-C8-R435Q cells as compared to HeLa-CD95-C8-WT cells (Fig. 3A, B; Supplementary Fig. 4A, B). Specifically, in these experiments, cells were stimulated with 500 ng/mL CD95L and caspase-8 and caspase-3/7 activities were monitored for one, two, three and four hours (Fig. 3A, B). In the case of HeLa-CD95-C8-R233H and HeLa-CD95-C8-R435Q cells, only a slight increase in caspase activity was observed compared to HeLa-CD95-C8-WT cells (Fig. 3A, B). It should be noted that the caspase-8 and caspase-3/7 activity assays are based on the cleavage of a short peptide-based substrate. However, these substrates are known to lack absolute specificity and to be cleaved not only by the above-mentioned caspases but also by the other caspases [38]. Therefore, these assays may actually indicate activation of the other caspases in addition to the one which activity is being tested, e.g., caspase-8 or caspase-3/7.
Fig. 3: Procaspase-8 mutations at the putative methylation sites inhibit CD95L-induced caspase activity and apoptosis.
A, B HeLa-CD95-C8-KO cells, which were transfected with pcDNA3 (vector), procaspase-8a-R233H-pcDNA3 (R233H), procaspase-8a-R435Q-pcDNA3 (R435Q) and procaspase-8a-WT-pcDNA3 (WT) or non-transfected (KO), were treated with the indicated amounts of CD95L for the indicated time intervals. Caspase-8 and -3 activities were determined by Caspase-Glo®8 Assay (A) and Caspase-Glo®3/7 Assay (B), respectively. Western Blot controls of transfection efficiency are shown in the lower part of the panels. C The HeLa-CD95-C8-KO cells were transfected as described before. This was followed by treatment with 500 ng/mL CD95L (+) for one hour and Western Blot analysis with the indicated antibodies were performed. One representative Western Blot out of three independent experiments is shown. D–F The HeLa-CD95-C8-KO cells were transfected as described before. Cells were treated with the indicated amounts of CD95L for indicated time intervals. Representative Western Blot analysis of transfection efficiency is shown in the lower part of the panels. D Cell viability was captured by measuring ATP levels using CellTiter-Glo® 2.0 Cell Viability Assay. E Metabolic activity was captured by measuring metabolic level using RealTime-Glo™ MT cell Viability Assay. F LDH release was measured by LDH-Glo® Cytotoxicity Assay. G, H HeLa-CD95-C8-KO cells were transfected as described above. After 24 hours, the cells were treated with indicated amounts of CD95L for 6 hours. Cells were stained by Annexin-V-FITC (An) and Propidium Iodide (PI) and then analysed by Imaging Flow Cytometry. G The amounts of viable cells (gated negative cells) after CD95L treatment are shown. H The amounts of An+ single-positive and An + /PI+ double positive cells after CD95L treatment are shown. I Representative images of viable (negative), An+ single-positive, An + /PI + positive (double-positive) cells. J The representative Western Blot analysis of transfection efficiency is shown. A, B, D–H Mean and standard deviation are shown for three independent experiments. Statistical analysis was carried out by ordinary ONE-WAY ANOVA with subsequent Tukey-test (** significant; p < 0.01; *** significant; p < 0.001; **** significant; p < 0.0001). s.e. short exposure, l.e. long exposure, Bf bright field, An, Annexin V-FITC, PI Propidium Iodide.
These results were consistent with the analysis of procaspase-8 and procaspase-3 processing by Western Blot in HeLa-CD95-C8-R233H and HeLa-CD95-C8-R435Q in comparison to HeLa-CD95-C8-WT cells (Fig. 3C, Supplementary Fig. 4C). The delayed rate of procaspase-8 and –3 cleavage was observed in HeLa-CD95-C8-R233H and HeLa-CD95-C8-R435Q compared to HeLa-CD95-C8-WT cells.
The decrease in caspase activity was consistent with the results of the analysis of the CD95L-induced loss of cell viability of HeLa-CD95-C8-R233H and HeLa-CD95-C8-R435Q compared to HeLa-CD95-C8-WT cells. The latter was monitored by assessing the total ATP levels and measuring cellular metabolism (Fig. 3D, E; Supplementary Fig. 4D). In these experiments, the introduction of the R233H and R435Q mutations into procaspase-8a resulted in an inhibitory effect on the CD95L-induced loss of cell viability. This was observed for both short-term and long-term CD95L treatment of HeLa-CD95-C8-R233H and HeLa-CD95-C8-R435Q cells by measuring total ATP levels and for the long-term treatment in the assays assessing cellular metabolism (Fig. 3D, E; Supplementary Fig. 4D).
The measurement of CD95L-induced LDH release also supported that the incorporation of the R233H and R435Q mutations into procaspase-8a inhibited CD95L-induced cell death in both short-term and long-term treatments, whereas the time-dependent increase in the CD95L-induced cytotoxicity was observed in HeLa-CD95-C8-WT cells (Fig. 3F). Finally, the analysis of the CD95L-induced apoptotic cell death using Annexin-V (An)/propidium iodide (PI) staining also showed that the introduction of the R233H and R435Q mutations into procaspase-8a blocked the CD95L-induced cell death (Fig. 3G–J, Supplementary Fig. 4E). Taken together, these experiments have demonstrated that the introduction of mutations at putative procaspase-8a methylation sites R233 and R435 leads to the inhibitory effects on CD95L-induced apoptosis.
Procaspase-8 may undergo symmetric di-methylationPRMT5 is able to methylate its substrates in two ways, via mono-methylation and symmetric di-methylation [26] (Fig. 4A). To explore whether procaspase-8 undergoes symmetric di-methylation we have performed IPs with the antibodies that recognize symmetric di-methylated proteins (SYM-10 antibodies). The SYM-10-IP should pulldown the proteins that undergo symmetric di-methylation. This IP was carried out from CD95L-stimulated HeLa-CD95-C8-KO cells transfected with procaspase-8a-WT, R233H, R435Q or vector control, e.g. HeLa-CD95-C8-WT, HeLa-CD95-C8-R233H, HeLa-CD95-C8-R435Q, or HeLa-CD95-C8-KO cells, respectively (Fig. 4B). The Western Blot analysis of SYM-10-IPs has revealed the presence of procaspase-8a-WT in SYM-10-IPs upon CD95L stimulation. However, no procaspase-8a-R233H or procaspase-8a-R435Q was detected in SYM-10-IPs (Fig. 4B). Furthermore, the p43 and p18 cleavage products of procaspase-8a were detected in the SYM-10-IP indicating that these two cleavage products can also undergo symmetric di-methylation. These results suggest a potential symmetric di-methylation of procaspase-8.
Fig. 4: Procaspase-8 may undergo symmetric di-methylation.
A Three types of arginine methylation are shown schematically. B HeLa-CD95-C8-KO cells transfected with pcDNA3 (vector), procaspase-8a-R233H-pcDNA3 (R233H), procaspase-8a-R435Q-pcDNA3 (R435Q) and procaspase-8a-WT-pcDNA3 (WT) as well as non-transfected (KO) were treated with 1000 ng/mL of CD95L (+) for one hour (h). This was followed by SYM-10-IP with subsequent Western Blot analysis. In these experiments, immunoprecipitation has been carried out without immobilization, which we designate as IP. Lysates (Input), IPs and bead-only control (B) are shown. One representative Western Blot analysis out of three independent experiments is shown. C HeLa-CD95-C8-KO cells transfected with pcDNA3 (vector), and procaspase-8a-WT-pcDNA3 (WT) as well as non-transfected (KO) were treated with 1000 ng/mL of CD95L (+) for one hour in the presence or absence of EPZ inhibitor. This was followed by SYM-10-IP with subsequent Western Blot analysis. Lysates (Input), IPs and bead-only control (B) are shown. One representative Western Blot analysis out of three independent experiments is shown. IP immunoprecipitation, B beads-only pulldown, s.e. short exposure, l.e. long exposure, v vector. B, C Non-specific signals are marked with *. The signal for the heavy chain of antibody used for IP is marked with IgGH.
EPZ (EPZ015666) is a specific inhibitor of PRMT5. Next, HeLa-CD95-C8-WT cells were co-treated with CD95L and EPZ. The SYM-10-IP from these cells showed that the addition of the EPZ inhibitor reduced the levels of procaspase-8a and its cleavage products p43 and p18 in the SYM-10-IP (Fig. 4C). This indicates a decrease in di-methylated procaspase-8 upon administration of EPZ, supporting the role of PRMT5 in the methylation of procaspase-8.
To provide further evidence for symmetric di-methylation, procaspase-8-IP from SKW 6.4 cells was performed with the addition of SDS and heating of the lysates to 95 °C, i.e. a so-called ‘denaturing’ IP of procaspase-8 (Den-C8-IP). The addition of SDS to Den-C8-IP led to the immunoprecipitation of procaspase-8 alone without any core DISC components such as FADD (Fig. 5A). Moreover, the Den-C8-IPs with subsequent Western Blot analysis using SYM-10 antibodies have provided further evidence for the possible symmetric di-methylation of procaspase-8. Indeed, the bands of the size of the caspase-8 cleavage products p43/p41 and p18 were detected in the Den-C8-IP from CD95L-stimulated SKW 6.4 cells using anti-SYM -10 Western Blot (Fig. 5A). As highlighted above, IPs performed under denaturing conditions contain only the bait, which in this experiment will be caspase-8. Naturally, PTM-modified caspase-8 isoforms will also be present in this pulldown. This gives further support for the symmetric di-methylation of procaspase-8 since anti-SYM-10 Western Blot analysis revealed the putative caspase-8 signals in the Den-C8-IPs.
Fig. 5: Caspase-8 activity is detected in SYM-10-IP.
A, B SKW 6.4 cells were treated with the indicated concentrations of CD95L for the indicated time intervals. A Denaturing (Den) Caspase-8-IP (Den-C8-IP) was analysed with anti-SYM-10 and anti-caspase-8 Western Blot. The position of p43/p41 and p18 bands detected with SYM-10 antibodies are indicated by arrows. B SYM-10-IP has been carried out. IPs and Lysate control (Input) were analysed by Western Blot with indicated antibodies. CSN3-IP was used as isotype control IP. One representative experiment out of three is shown. Heavy chain of antibody is marked with IgGH. C SYM-10-IP has been carried out. IPs and Lysate control (Input) were analysed by Western Blot with indicated antibodies. One representative experiment out of three is shown. D Caspase-8 activity at SYM-10-IPs shown in (C) was determined using the IPs. Mean and standard deviation are shown for three independent experiments. Statistical analysis was carried out by unpaired student t-test (** significant; p < 0.01). IP immunoprecipitation, B beads-only pulldown, s.e. short exposure, l.e. long exposure, IgGH Heavy chain of immunoprecipitating antibody, IgGL light chain of immunoprecipitating antibody.
Since PRMT5 methylates its substrates via mono-methylation and symmetric di-methylation, we also investigated if procaspase-8 might undergo mono-methylation. However, the analysis of Den-C8-IP by Western Blot with anti-mono-methylation (MMA) antibodies did not reveal any signals corresponding to the molecular weight of procaspase-8 or its cleavage products (Supplementary Fig. 5A). This observation was further supported with the IP from SKW 6.4 cells without the addition of SDS using anti-MMA antibodies. This also did not result in immunoprecipitation of procaspase-8 (Supplementary Fig. 5B). Taken together, these experiments did not reveal any mono-methylation of procaspase-8.
Pharmacological inhibition of methylation reduces CD95L-induced caspase activityIt should be noted that in HeLa-CD95-C8-KO cells transfected with procaspase-8a-WT, procaspase-8a cleavage upon CD95L stimulation, was strongly decreased upon administration of EPZ (Fig. 4B). In line with the decrease in procaspase-8a processing, the CD95L-induced cleavage of PARP1, was also downmodulated upon administration of EPZ (Fig. 4B). These results strongly suggest that the catalytic activity of caspase-8 and effector caspases is inhibited upon pharmacological inhibition of PRMT5.
To further investigate the link between caspase-8 activity and symmetric di-methylation, SYM-10-IPs were carried out from CD95L-stimulated and non-stimulated SKW 6.4 cells with the following analysis of caspase-8 activity. SYM-10-IPs from CD95L-stimulated cells contained the procaspase-8 cleavage products p43/p41 and p18 (Fig. 5B, C). Furthermore, caspase-8 activity can be detected in these IPs upon CD95L stimulation (Fig. 5D). The latter provides further support that symmetrically di-methylated caspase-8 has a catalytic activity.
Consistent with the results obtained in HeLa-CD95 cells (Fig. 4B), the rate of procaspase-8a/b processing in CD95L-treated SKW 6.4 cells was also decreased upon EPZ administration, as evidenced by the diminished amounts of p43/p41 in the total cellular lysates (Fig. 6A). This analysis further indicates that the inhibition of methylation by means of the addition of EPZ decreases the rate of procaspase-8 activation.
Fig. 6: Pharmacological inhibition of PRMT5 blocks CD95L-induced caspase cascade.
A, B SKW 6.4 cells were treated with 250 ng/mL of CD95L, zVAD-fmk and EPZ for the indicated time intervals with subsequent Western Blot analysis. One representative experiment out of three independent ones is shown. C–H BT549 cells (C, F), U67 (D, G) and HT29 (E, H) were treated with CD95L for two hours (C–E) or four hours (F–H). Caspase-8 and -3/7 activities were determined by Caspase-Glo®8 Assay (C–E) and Caspase-Glo®3/7 Assay (F–H), respectively. A–H EPZ pre-treatment was performed for two hours before CD95L treatment and zVAD-fmk pre-treatment was performed for one hour before CD95L stimulation. Statistical analysis was carried out by ordinary ONE-WAY ANOVA with Tukey-test (ns not significant; p > 0.05, * significant; p < 0.05, ** significant; p < 0.01; *** significant; p < 0.001; **** significant; p < 0.0001). s.e. short exposure.
In line with the decrease of procaspase-8 activity, the analysis of caspase-3 and PARP1 cleavage by Western Blot in CD95L-treated SKW 6.4 cells showed that EPZ delayed the cleavage of the effector caspase-3 as well as its substrate PARP1 (Fig. 6B). These effects were consistent with the inhibitory effects of AMI-5, which is the pan-methylation inhibitor of protein arginine methyltransferases. In particular, AMI-5 inhibited the activity of effector caspases-3 and -7 in SKW 6.4 cells (Supplementary Fig. 6A). Moreover, administration of EPZ in breast carcinoma BT549, glioblastoma U87 and colon carcinoma HT29 cells led to the decrease in CD95L-induced caspase-8 (Fig. 6C–E) and caspase-3/7 activities (Fig. 6F–H).
Collectively, these results show that the pharmacological inhibition of PRMT5 leads to the inhibitory effects on the caspase cascade.
Pharmacological inhibition of methylation decreases CD95L-mediated apoptosis upon short-term stimulationThe next step was to investigate the impact of pharmacological inhibition of methylation on extrinsic cell death pathways. Since caspase-8 is a negative regulator of necroptosis, we next examined whether administration of EPZ could affect CD95L-mediated necroptosis induction, which was induced by co-treatment of CD95L, IAP inhibitor BV6 and zVAD-fmk in HT29 cells. Western Blot analysis showed phosphorylation of MLKL and RIPK1 upon CD95L/BV6/zVAD-fmk treatment, but no effect on this phosphorylation was detected upon co-administration of EPZ, e.g. upon CD95L/BV6/zVAD-fmk/EPZ compared to CD95L/BV6/zVAD-fmk treatment (Fig. 7A). The latter may be related to the fact that EPZ inhibits caspase-8 activity, which is already inhibited by zVAD-fmk administration in these experiments. Importantly, however, treatment with CD95L, EPZ and BV6 alone, without the addition of zVAD-fmk, induced the phosphorylation of MLKL and RIPK1, supporting the hypothesis that EPZ inhibits caspase-8 activity and thereby may also promote the induction of necroptosis (Fig. 7B).
Fig. 7: Pharmacological and genetic downmodulation of PRMT5 activity reduces cell viability upon long-term treatment.
A, B HT29 cells were treated with the indicated concentrations of CD95L, EPZ, zVAD-fmk and BV6 for indicated time intervals. Necroptosis markers were evaluated by Western Blot with the indicated antibodies. C HT29 cells were treated with the indicated concentrations of EPZ and CD95L for the indicated time intervals. Cell viability was captured by measuring ATP levels using CellTiter-Glo® 2.0 Cell Viability Assay. C–F HeLa CD95 cells were transfected by siRNA of PRMT5 and control siRNA. Western Blot analysis (D) and Quantification (E) of transfection efficiency are shown. F Effects of the PRMT5 KD on cell viability was captured by measuring ATP levels using CellTiter-Glo® 2.0 Cell Viability Assay and normalized against untransfected cells. C, E, F Mean and standard deviation are shown for three independent experiments. Statistical analysis was carried out by ordinary ONE-WAY ANOVA and followed Tukey-test (** significant; p < 0.01; *** significant; p < 0.001; **** significant; p < 0.0001).
Next, we aimed to analyse the effect of inhibiting caspase-8 methylation on long-term co-treatment with CD95L. To this end, we first tested the viability of the cells upon EPZ treatment alone (Fig. 7C). However, this analysis revealed that long-term co-treatment with EPZ alone severely reduced cell viability, precluding further analysis of the effects of pharmacological inhibition of PRMT5 on caspase-8 activity upon long-term CD95L/EPZ stimulation. This is consistent with previous reports on the essential role of PRMT5 in key cellular processes [39]. In line with these results, siRNA silencing of PRMT5 also resulted in a significant loss of cell viability as early as 24 hours after transfection (Fig. 7D–F, Supplementary Fig. 6B, C). These results precluded further investigation of the effects of genetic or pharmacological inhibition of PRMT5 on CD95L-mediated cell death upon long-term treatment.
Despite the effects observed upon long-term treatment, we analysed the effects of EPZ on CD95L-mediated cell death upon short-term stimulation, considering that under these conditions, PRMT5 inhibition does not yet affect cell viability. Indeed, when HeLa-CD95 cells were stimulated with 250 ng/ml CD95L for 6 hours, the addition of EPZ blocked the rate of cell death as measured by imaging flow cytometry (Fig. 8A–C). The latter results are consistent with the data obtained above and show that inhibition of PRMT5 upon short-term treatment leads to the inhibitory effects on caspase-8 activation, caspase cascade and subsequent apoptosis.
Fig. 8: Pharmacological inhibition of PRMT5 blocks CD95L-induced apoptosis.
A–C HeLa-CD95 cells were treated with indicated amounts of CD95L for 6 hours. Cells were stained by Annexin-V-FITC (An) and Propidium Iodide (PI) and then analysed by Imaging Flow Cytometry. A The amounts of viable cells (negative cells) after CD95L treatment are shown. B The amounts of An+ single-positive and An + /PI+ double-positive cells after CD95L treatment are shown. C Representative images of viable (negative), An + single-positive, An + /PI + positive (double-positive) cells. A, B Statistical analysis was carried out by ordinary ONE-WAY ANOVA with Tukey-test (ns not significant; p > 0.05, * significant; p < 0.05). Bf bright field, An Annexin V-FITC, PI Propidium Iodide. D The proposed role of caspase-8 methylation in the CD95 apoptotic cascade. The assembly of the CD95 DISC consisting of CD95, FADD, procaspase-8 and c-FLIP is shown leading to caspase-8 activation. The putative caspase-8 methylation sites, arginines 233 and 435, are shown as green spheres. The proposed effects of PRMT5 on caspase-8 activation are shown schematically.
Comments (0)