
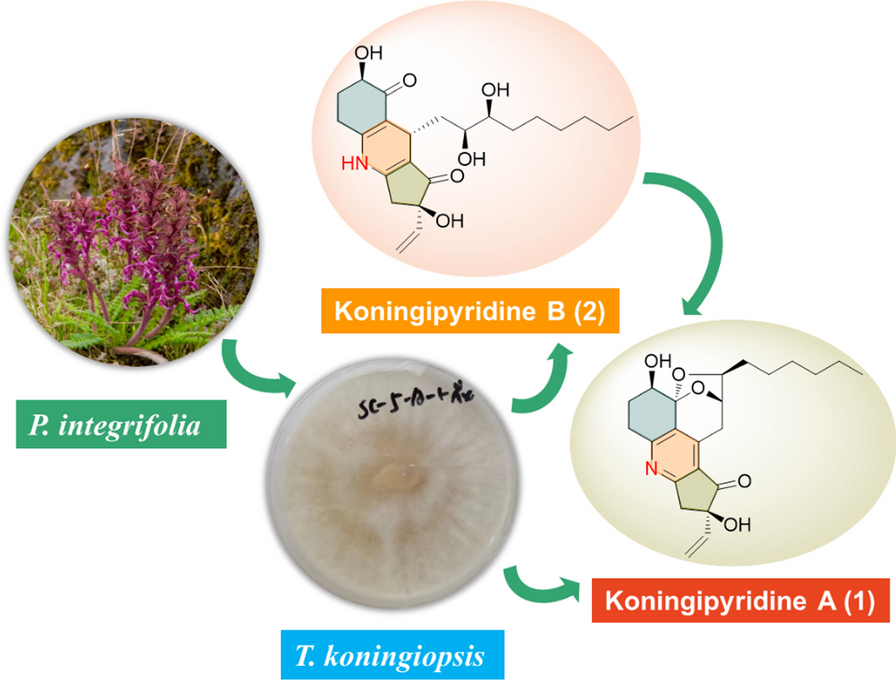
記住我
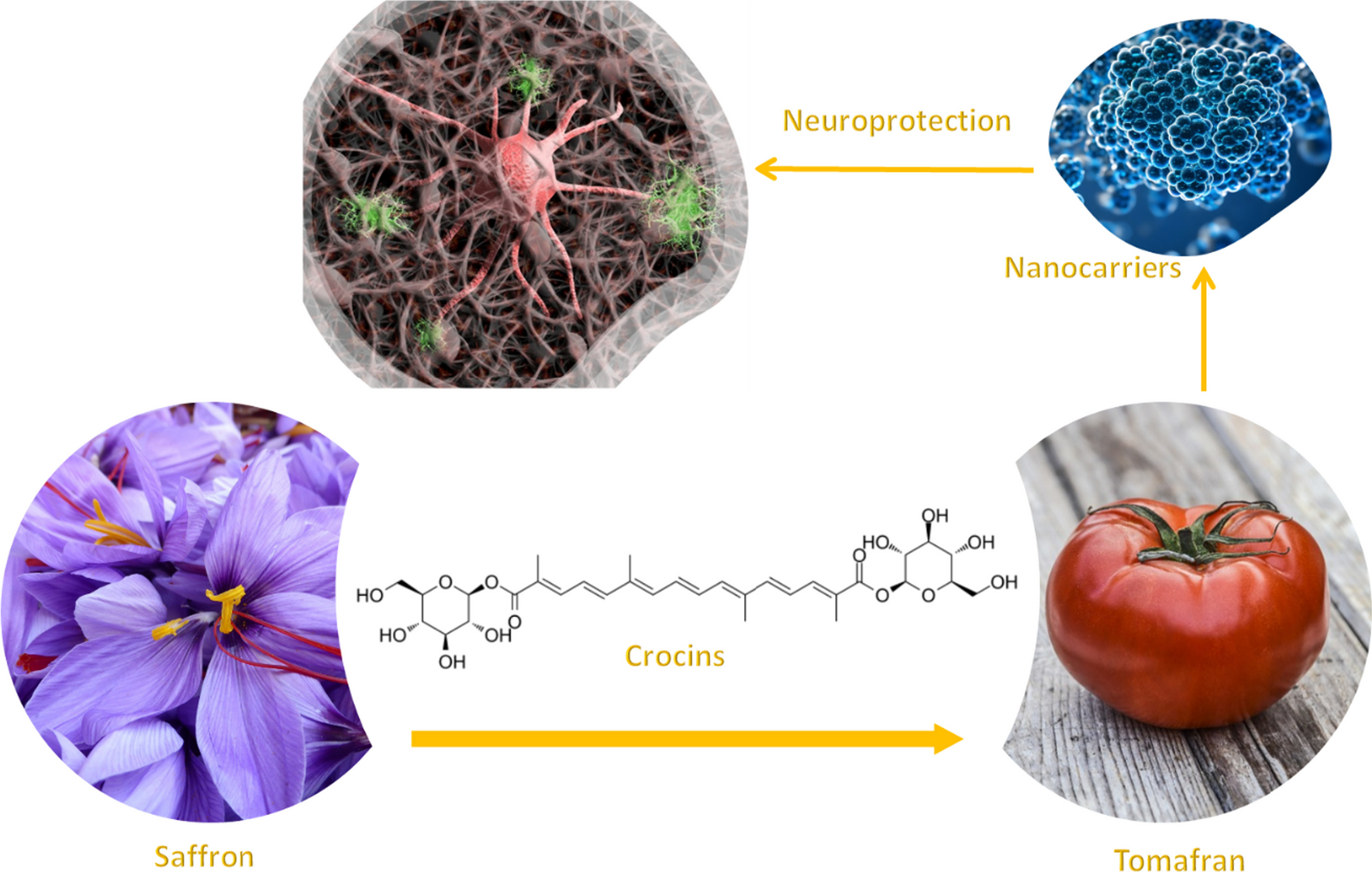
With the aim to evaluate the potential neuroprotective activity, different extracts rich in crocins have been evaluated. Tomato fruits from wild type and bio-engineered tomato were examined for the presence of saffron apocarotenoids (crocetin, crocins, picrocrocin, and safranal) after the seeds had been removed. The tomatoes from both lines were crushed and the serum juice was collected and lyophilized. As a control, a water extract from saffron dried stigma was used. Crocins were quantify by HPLD-DAD, and extracts from saffron and Tomafran were diluted to a concentration of 100 µM of crocins while the tomato extract was adjusted to the corresponding amount of Tomafran by weight. Then, these extracts were studied in a cellular model of early staged of Alzheimer’s disease (AD) characterized by high phosphorylation of microtubule-associated protein tau. It is well known that treatment of neuroblastoma SH-SY5Y cells with okadaic acid (OA), a protein phosphatase inhibitor, produces neurotoxicity by high molecular weight phospho-tau species [18], being the cellular model here used. Firstly, and to check that the extracts were not toxic for the cells, cell viability after extracts treatment was measured using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide) method. The results showed that both tomato extracts did not reduce cell viability, when a 100 µM of crocin in Tomafran extract was used, however saffron extract reduced cell viability by 27% (Fig. 1A).
Fig. 1
A Percentage of cellular viability of SH-SY5Y using tomato, Tomafran and saffron extracts. B Percentage of cellular viability of SH-SY5Y after OA treatment, using tomato, Tomafran and saffron extracts. (*P < 0.05, **P < 0.01, ***P < 0.001). (One-way ANOVA with a post hoc Tukey test)
Treatment with 70 µM of OA was used to inhibit the activity of protein phosphatases in undifferentiated neuroblastoma SH-SY5Y cells that express tau endogenously, decreasing the cell viability of about 43%. Only Tomafran extract was able to reverse the cell death significantly, pointing out its neuroprotective effect against this toxin (Fig. 1B), and showing its potential as herbal treatment, food supplement or nutraceutical for AD, being safer than saffron extracts.
2.2 Prediction of CNS penetration of the different extracts rich in crocinsAs previously mentioned in the introduction part, one of the main obstacles for the treatment of the diseases of the central nervous system (CNS) is the drug’s penetration into the brain at therapeutic concentrations. Most CNS drugs cross the BBB by transcellular passive diffusion, due to the tight junction structure and limited transport pathways. Parallel artificial membrane permeability assay (PAMPA) is a high throughput technique developed to predict passive permeability through biological membranes. In order to explore the capacity of tomato, saffron and Tomafran extracts to penetrate into the brain, a PAMPA-BBB method employing a brain lipid porcine membrane was carried out [19]. The in vitro permeabilities (Pe) of ten commercial drugs for human therapy through lipid membrane extract together with tomato, saffron and Tomafran extracts were determined and described in Table 1. An assay validation was made comparing the reported permeabilities values of commercial drugs with the experimental data obtained employing this methodology. A good correlation between experimental-described values was obtained Pe (exptl) = 1.1108 (bibl)—0.9618 (R2 = 0.961) (Fig. 2). From this equation and following the pattern established in the literature for BBB permeation prediction [20], we could classify compounds as CNS + when they present a permeability > 4.04 × 10–6 cm s−1. Based on these results we can consider that tomato, saffron and Tomafran extracts are not able to cross the BBB by passive permeation (Table 1).
Table 1 Permeability (Pe 10–6 cm s−1) in the PAMPA-BBB assay for 10 commercial drugs (used in the experiment validation) and tomato, saffron and Tomafran extracts with their predictive penetration in the CNSFig. 2
Linear correlation between experimental and reported permeability of commercial drugs using the PAMPA-BBB assay
However, previous reported results showed that crocins cross the BBB by passive permeation at higher time-points [21]. Furthermore, crocins are digested in the colon to produce deglucosylated trans-crocetin, which is then rapidly and extensively absorbed beyond the intestinal barrier via passive transcellular diffusion [22]. In this sense, we decided to test in the PAMPA-BBB assay, Tomafran and previously digested Tomafran extract for 4, 8, 12, 16, 20 and 24 h. Again, an assay validation was made comparing the reported permeabilities values of commercial drugs with the experimental data obtained employing this methodology. A good correlation between experimental-described values with the corresponding equation was obtained (Table 2).
Table 2 Obtained correlation (r2) for equations of each time-point experimentAs previously described, compounds were classified as CNS +/±/− depending on the obtained equation. The results indicated that Tomafran extract is not able to cross the BBB at any of the tested times. Meanwhile, the previously digested extract of Tomafran was able to penetrate the BBB within 20 h with a Pe (10–6 cm s−1) of 5.20 which may indicate that trans-crocetin might permeates BBB with a slow but constant velocity over a 24 h period in agreement with reported studies (Table 3). Anyhow, better strategies for brain delivery would be desirable.
Table 3 Tomafran extract and digested Tomafran with their predictive penetration in the CNS at 4, 8, 12, 16, 20 and 24 h2.3 Tomafran extract encapsulation into chitosan nanoparticles and in vitro cellular studiesIn order to improve the delivery of Tomafran extracts into the CNS, we decided to encapsulate the extract in chitosan nanoparticles. Polymeric nanocarriers (NCs) based on low MW chitosan are potential candidates for the non-invasive conveyance of therapies via the BBB [23, 24] having many benefits, such as low toxicity, biodegradability, biocompatibility, and mucoadhesivity. Moreover, chitosan has other advantages such as the simplicity of functionalization due to functional groups on its structure, which further enhances the capabilities of chitosan-based NCs. Therefore, to improve the velocity of permeability we decided to encapsule Tomafran extracts (TFE) into chitosan (CH) nanoparticles.
Different nanoparticles formulations were obtained using two CH:TFE ratios formulations 5:1 and 1:1. To determine the correct proportions of raw material and plant extract, photonic spectroscopy to measure of average size, surface Z potential, and polydispersity index (PDI) of each ratio formulations was used. The results presented in Table 4, showed nanoparticles average size ranges from 49 to 106 nm of diameter; showing an increase of the size of the nano vehicle related to the TFE encapsulation. However, nanoparticles (NPs) with the ratio 1:1 and 5:1 shown similar size in agreement with previous works reporting that plant extracts chitosan nanoparticles did not displayed a significative increase of average size due to a higher molecular weight of raw material [25]. Polydispersity (PDI) index was measured to know the homogeneity of nano-formulations, Table 4 shows the decrease of PDI from 0.4 to 0.3 in NPCH (chitosan nanoparticles) and NPCH-TFE (chitosan nanoparticles containing Tomafran extract) formulations respectively. The incorporation of glycosylated apocarotenoids could provide a stabilizing effect due to the alkyl glycosidic structure conferring non-ionic surfactant properties [26].
Table 4 Average size, polydispersity (PDI), Z-potential, encapsulation efficiency (EE) and loading capacity (LC) for NPs characterizationSurface Z potential is a predictive value of stability and quality of nano-formulations, where values near to + 30 mV and − 30 mV indicates a stable colloid suspension [27]. Furthermore, the surface charge is indicative of the electrochemical properties of the nanoparticles, since the type of charge provides different physicochemical and biological properties, besides different approaches to surface chemical modifications of the nanoparticles. The results showed in Table 4 presented a positive surface charge with an interval ranging from + 31.4 to + 24.2 mV in NPCH and NPCH-TFE 1:1 respectively. Nanoparticles showed a characteristic positive surface Z potential related to the use of chitosan raw material and non-ionic surfactants with suitable values close to + 30 mV indicating a well stabilized suspension. The decrease of surface zeta potential has been related to the increase of TFE into chitosan nanoparticles in agreement with previous works where the increase of active molecule in chitosan nanoparticles decrease the surface charge [28].
Finally, transmission electron microscope (TEM) was used to determine the surface and morphological characteristic of chitosan nanoparticles [29]. The images in Fig. 3 corresponding to two different TEM micrograph magnifications showing in both cases spherical shaped nanoparticles with some aggregates. The size of the nanoparticles is distributed between 10 and 30 nm and differs from the size obtained by Dynamic Light Scattering (DLS), confirming the swelling and aggregation effect observed previously in other works with chitosan nanoparticles due to self-aggregation among individual nanoparticles and or/swelling of the chitosan layer surrounding the nanoparticle in aqueous media. Because of these phenomena, the size and PDI measurements in DLS show a higher size than the TEM technique [30].
Fig. 3
A NPCH-TFE image with 100 nm scale bar. B NPCH-TFE image with 50 nm scale bar
Once the NPCH-TFE were obtained and physico-chemically characterized, different concentration of encapsulated Tomafran extracts ranging from 100 to 6.25 μM were assayed to check SH-SY5Y cell viability. The formulations NPCH-TFE 5:1 and NPCH-TFE 1:1 have been assessed using SH-SY5Y cells in the presence of okadaic acid (OA), and the results obtained were comparable among them. Only data from NPCH-TFE 1:1 are presented. As shown in Fig. 4A, NPCH-TFE 1:1 containing concentration from 50 to 100 μM were toxic to SH-SY5Y cell and reduce the cell viability by an average of 50% while NPCH-TFE 1:1 containing lower concentration of crocins were harmless to cells. Therefore, neuroprotection assays against OA at doses ≤ 25 µM crocins were carried out. As shown in Fig. 4B, the neuroprotective activity of NPCH-TFE 1:1 at 25 µM is statistically significant.
Fig. 4
A Percentage of cellular viability of SH-SY5Y using NPCH-TFE containing different concentration of crocins. B Percentage of cellular viability of SH-SY5Y after OA treatment, using different crocin’s doses of NPCH-TFE. Asterisks denote significant differences between OA group and treated groups (*P < 0.05, **P < 0.01, ***P < 0.001). (One-way ANOVA with a post hoc Tukey test)
Finally, the artificial membrane permeability test (PAMPA) was carried out under the same previously described conditions. The new formulations NPCH-TFE 5:1 and NPCH-TFE 1:1 were able to cross the barrier by passive diffusion with a Pe (10–6 cm s−1) of 14.8 ± 1.8 and 14.4 ± 3.4, respectively. Compared to the TFE solution, passive penetration of NPCH-TFE was significantly improved. Neither TFE membrane retention nor binding to the filter's surface occurred. Thus, the data show that the NPCH-TFE formulations successfully ameliorated passive permeation trough the BBB.
2.4 Characterization of tomato and Tomafran derivate exosomesIn addition to NPCH-TFE, we decide to also study the neuroprotective capacity of tomato and Tomafran exosomes. Exosomes originating from particular plant sources, such as ginseng or green tea, have been the subject of some studies looking into their therapeutic potential in Alzheimer’s disease animal models. These investigations have shown encouraging findings, indicating that plant exosomes may have neuroprotective qualities and may be employed as therapeutics [31]. For all the above reasons, we aimed to purify exosomes from tomato wild type and Tomafran to find out whether they have a potential neuroprotective activity. In this work, we collected and screened two plant-derived exosome from a wild-type tomato and transgenic Tomafran in order to offer a new alternative to treat CNS disorders based on these natural extra vesicles. Before the in vitro cytotoxicity evaluation, we first characterized the different isolation of exosomes from tomato and Tomafran using TEM (Fig. 5) and nanotraking analysis (NTA) (Fig. 6).
Fig. 5
Scanning electron micrographs of fixed and dehydrated exosomes from wild type tomato (A) and Tomafran (B)
Fig. 6
Quantification of tomato money maker (A) and Tomafran (B) vesicle concentration by nanoparticle tracking analysis (NTA)
To have a closer look at the morphology of these vesicles, TEM analysis confirmed the presence of vesicular structures. Preparations of exosomes with a final protein concentration of 1.9 mg/mL in Tomafran and 3.7 mg/mL in tomato were used. To have a closer look at the morphology of these vesicles, we performed a TEM analysis that confirmed the presence of vesicular structures. Both exosomes preparations have an average size of 140 nm (Fig. 5) with a typical cup-shaped morphology. Most exosomes are spherical, but sample fixation and dehydration result in cup-shaped morphology observed by SEM and TEM [32].
NTA is a widely technique that permits the determination of both the size distribution and relative concentration of microvesicles, including exosomes, in the supernatants of cultured cells and biological fluids [33]. The NTA of purified exosomes are shown in Fig. 6. In both cases, the exosomes displayed a wide size distribution of particles with higher number of vesicles less than 200 nm achieving a maximum concentration of particles with 137 nm and second minor population with 188 nm in the case of wild type tomato exosomes. Meanwhile, the exosomes population in transgenic Tomafran showed three main populations of nanovesicles with 65 nm, 169 nm and 217 nm. The distribution observed in the case of tomato exosomes was similar to the profile obtained recently by Kilasoniya and co-workers where the exosomes obtained from tomato juice showed a main population of PELNs ranging from 140 to 170 nm [34]. Another recent work described a similar pattern in the size distribution of exosomes obtained from tomato confirming the main population with exosomes with 180 nm [35].
Neuroprotection studies were done with different concentrations of Tomafran exosomes as shown in Fig. 6. Results shown that exosomes from tomato wild type were not able to protect the SH-SY5Y cells from the damage caused by OA while exosomes from Tomafran at doses of 25 and 50 μg/mL protect the cells significatively (Fig. 7). This effect may be due to the presence of crocins which exert its strong antioxidant activity by reducing oxidative stress and inflammation.
Fig. 7
Percentage of cellular viability of SH-SY5Y after OA treatment, using (A) exosomes from Tomato wild type (B) Tomafran. Asterisks denote significant differences between OA group and treated groups (*P < 0.05, **P < 0.01, ***P < 0.001). (One-way ANOVA with a post hoc Tukey test)
The use of plant exosomes in a therapeutic setting, either by leveraging known therapeutic capabilities of plants or by packing plant exosomes with a therapeutic cargo, is a recurring future prospect.
留言 (0)