
記住我
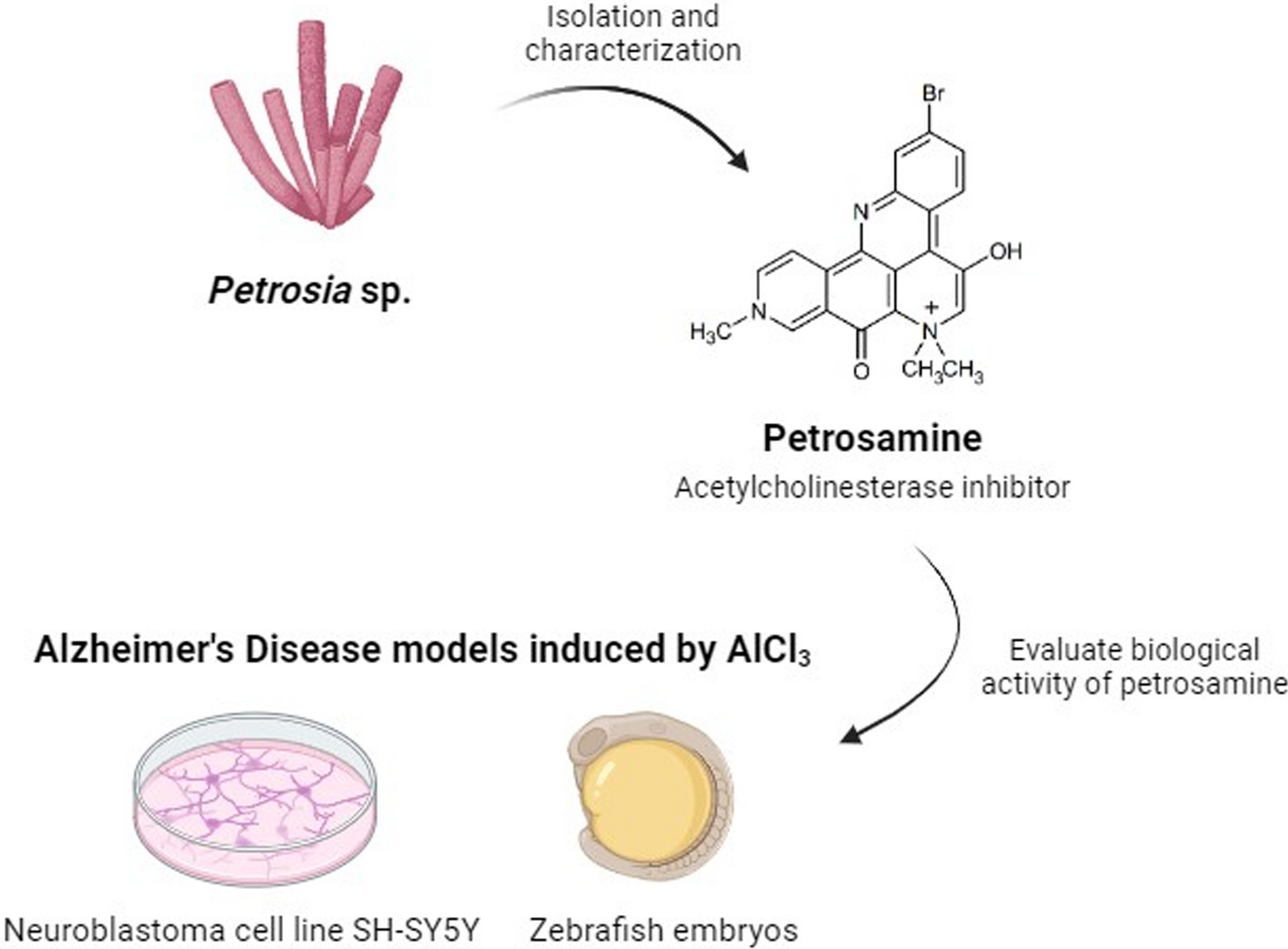
The newly collected sponge was immersed in ethanol and stored in a freezer. Two portions of approximately 500 g of the sponge material were cut into small pieces and extracted with MeOH (2 × 600 mL) under an ultrasonication bath. The extracts were combined and the solvent evaporated at reduced pressure (40 °C) to approximately 200 mL of a hydromethanol fraction which was partitioned with n-hexane, CH2Cl2, and EtOAc (3 × 200 mL of each solvent). The resulting fractions were dried with anhydrous sodium sulfate (Na2SO4) and concentrated under reduced pressure to yield fractions n-hexane (1.4 g) CH2Cl2 (0.7 g), and EtOAc (0.2 g). The CH2Cl2 fraction was subjected to a Sephadex LH-20 column and eluted with a binary mixture of CH2Cl2/MeOH 2:8. A subfraction (213 mg) enriched in a deep blue material was purified by HPLC using a phenyl-hexyl semi-preparative column and a gradient solvent system of H2O (0.01% TFA)/MeOH 60–100% by 15 min, flow rate 3.0 mL/min, temperature 35 °C, to yield a pure compound (45.5 mg, tR 10.6 min (Additional file 1: Fig. S1), which was unequivocally identified as petrosamine (Fig. 1) based on 1D and 2D NMR spectra.
Fig. 1
Chemical structure of petrosamine
2.1.1 Spectroscopic dataDark blue needles: mp 330–332 °C; UV-PDA (H2O/MeOH) lmax 286, 345, 589 nm. 1H NMR (300 MHz in MeOD) δH (ppm): 9.80 (1H, s, H-9), 9.45 (1H, d, J = 5.7 Hz, H-12), 9.13 (1H, d, J = 9.0 Hz, H-4), 9.05 (1H, d, J = 5.7 Hz, H-11), 8.29 (1H, br s, Hz, H-1), 7.78 (1H, br d, J = 9.0 Hz, H-3), 4.63 (2H, s, 2H-6), 4.67 (3H, s, Me-14), 3.93 (6H, s, Me-15 and Me-16). 13C NMR (75 MHz in MeOD) δC (ppm): 188.1 (C-5), 162.5 (C-8), 146.7 (C-9), 144.7 (C-13a), 144.1 (C-12b), 143.2 (C-11), 141.2 (C-12a), 136.7 (C-3), 133.7 (C-8a and C-1), 130.1 (C-12c), 127.6 (C-4), 124.4 (C-4a), 123.9 (C-12), 122.6 (C-2), 117.2 (C-4b), 116.3 (C-7a), 72.0 (C-6), 54.7/54.5 (C-15 and C-16), 49.3 (C-14) (Additional file 1: Figs. S2, S3).
2.2 Petrosamine reveals a good biocompatibility profileDespite all the efforts from the scientific community, disease-modifying treatments for AD are not yet available [15]. This neurodegenerative disease is among the pathologies with the lowest rate of drug development success, with 99% of drug candidates being discontinued after showing no clinical benefit [16]. Therefore, since the approval of galantamine for the treatment of AD patients, the search for new anticholinesterase alkaloids has escalated [11]. Petrosamine, a pyridoacridine alkaloid, is a natural compound isolated from marine sponges shown to be 6 times more potent than galanthamine in inhibiting AChE [17], with a AChE IC50 0.091 µM, but not yet tested systematically in vitro or in vivo [10] for its safety profile. Petrosamine’s biocompatibility was hereby tested to verify the best suitable concentration from a range of 0.01 to 0.15 mg/ml, using: (1) cytotoxicity in neuroblastoma cell line (SH-SY5Y); (2) fish embryotoxicity assay (FET) with zebrafish; and (3) hemolysis assay.
Cell viability was evaluated in SH-SY5Y cells by resazurin after exposure to petrosamine for 24 h (Fig. 1a). The percentage of cell survival for each sample was expressed and normalized against the life control (100% viability). The rate of viability slightly increased only with the lowest concentration of petrosamine (0.01 mg/ml) and then decreased with increasing concentration of this compound, being slightly lower than the control but without statistically significance, which attests to petrosamine being non cytotoxic in all tested concentrations. Interestingly, as pyridoacridine alkaloids are considered DNA binding molecules, their cytotoxicity screenings have mainly been focused in identifying those that kill cancer cells and not as much to guarantee their lack of cytotoxicity, particularly in the CNS [18].
Using the zebrafish embryotoxicity assay, recommended by the OECD for evaluation of acute toxicity of chemicals (OECD test guideline (TG) 236), the survival rate of the zebrafish embryos was tested until 120 h post fertilization (hpf), as observed in Fig. 1b. The four tested concentrations of petrosamine were added to the water from 72 hpf until 120 hpf, as the AlCl3 treatment was applied in the first 72 h of fish development [19]. The highest concentration of petrosamine was lethal to all embryos at 120 hpf, while the remaining concentrations supported a survival rate of 100% for all timepoints. Cesário et al., evaluated the cytotoxicity of three main isolated compounds (0.1, 0.5, and 1.0 mg/mL) from the marine sponge Aplysina fulva, on adult zebrafish, and showed that those concentrations tested were non-toxic, which goes accordingly to our results [20].
In order to determine the effects of petrosamine in erythrocytes, as intravenous application the most probable route of its administration, the hemolysis assay was performed. The determination of the hemolytic properties of petrosamine, in vitro, is an important step for a preliminary evaluation of interaction with the membrane of red blood cells to determine their safety, as it permits to understand their propensity to interact with cell membrane causing lysis. Results shown in Fig. 1c are normalized for the death control consisting of Triton 10 × at 10% (v/v). Based on all the results regarding its safety profile, the chosen concentration of petrosamine for subsequent studies on its capacity to counteract the AlCl3-induced neurotoxicity was 0.05 mg/ml.
2.3 Petrosamine counteracts aluminium-induced neurotoxicity in vitroThe scarce development of novel disease-modifying treatments and failed clinical trials are due to several factors, including premature translation of highly successful drugs in animal models that mirror only limited aspects of AD pathology to humans [21]. Consequently, there is a real necessity for model systems that can reproduce the pathological and biochemical characteristics of AD with the aim to allow adequate, fast preclinical screening of novel therapeutics [6]. In order to establish an AD model in vitro, cells from the neuroblastoma cell line SH-SY5Y were exposed to different concentrations of AlCl3 (50, 200 and 1000 µM) for 24 h. Metabolic viability was assessed with resazurin assay and the percentage of cell survival for each sample was expressed and normalized against the life control (100% viability). The resazurin assay showed that 50, 200 and 1000 µM petrosamine reduced cell viability by 14.0% (p < 0.05), 20.6% (p < 0.01) and 23.4% (p < 0.001), respectively (Fig. 2). In AD, there is a significant degree of apparent neuronal death in various areas of the brain [22] and AlCl3 properly reproduces this hallmark. A direct correlation between the increasing concentration of AlCl3 and the decrease in cell viability was observed. The concentration of 200 µM reduced cell viability to 79.3%, values very similar to those seen by Rizvi et al., that conducted a similar study (MTT assay) with 200 µM Al(mal)3 (74.5% cell viability) [23], p. 53]. Yang et al. have shown that the exposure of SH-SY5Y cells to aluminium ions in concentrations below 100 µM does not cause a significant reduction in cell viability and proliferation, which is consistent with the results obtained [24]. Thus, significant cell death could be observed at 1000 µM petrosamine, with 76.6% of cell viability.
Fig. 2
Biocompatibility of petrosamine. a Cytotoxicity of petrosamine in SH-SY5Y cell line after 24 h of incubation; b Survival rate of zebrafish embryos incubated with petrosamine from 72 to 120 hpf; c Evaluation of toxicity in erythrocytes incubated with petrosamine by hemolysis quantification
Using this new in vitro model, results demonstrated that petrosamine (0.05 mg/ml or 0,01 mM) is efficient in restoring cell viability affected by 24 h exposure to AlCl3, since this natural compound could restore cell viability levels to those of the life control, when compared to cells incubated with AlCl3 only.
Acridine orange (AO) / Propidium iodide (PI) assay was used to determine how AlCl3 induced neurotoxicity and to understand if the protection conferred by the petrosamine could translate into a reduction of apoptosis. While AO is a cell permeable green fluorescence probe, PI is a red fluorescent probe that only marks cells with compromised cell membranes and, consequently, stains only late apoptotic or necrotic cells. The double staining of the cells incubated with AlCl3 is presented in Fig. 3a to h. For this assay, two controls were performed, the life control (100% cell viability) and the death control (0% cell viability). In Fig. 3i, the percentage of viable cells, early apoptotic cells and late apoptotic/necrotic cells in each condition are reported. The number of cells allocated to each category was determined using ImageJ software. The AO/PI assay showed that 50, 200 and 1000 µM petrosamine led to 86.2% (p < 0.05), 79.8% (p < 0.01) and 57.3% (p < 0.0001) of viable cells, respectively. Thus, the number of early apoptotic and late apoptotic/necrotic cells increases with 24 h incubation with higher concentrations of AlCl3. As it is evident in Fig. 3, cells treated with petrosamine subsequently to AlCl3 incubation were mostly stained green, showing few apoptotic cells, while cells exposed to different concentrations of AlCl3 showed a significant portion of cells stained yellow and red, a marker of early apoptosis and apoptosis. In this assay, a reduction in apoptosis and concomitant restoration of cell viability are observed in the following conditions: 50 µM and petrosamine (95.8%), 200 µM and petrosamine (94.9%) and 1000 µM and petrosamine (94.0%).
Fig. 3
Evaluation of neuroprotection conferred by petrosamine in SH-SY5Y cell line exposed to AlCl3 for 24 h, measured by resazurin assay. Percentages are expressed relative to the life control (100% cell viability). Death control (30% DMSO). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 when compared to the life control
Lipid peroxidation occurs when ROS react with susceptible unsaturated lipids on the cell membrane, potentially causing irreversible damage and it is known that Al leads to a higher production of ROS, stimulating iron-induced membrane lipid peroxidation [12]. To determine the lipid peroxidation level in SH-SY5Y cells, the TBARS assay was performed, and results of each sample were normalized against life control (100%). As shown in Additional file 1Fig. S1, no statistically significant differences were identified in lipid peroxidation of cells submitted to treatment with AlCl3, meaning that the tested concentrations do not have a substantial impact over this hallmark. Nevertheless, a clear trend is verified with the percentage of lipid peroxidation increasing with higher concentrations of AlCl3. Neurons are very sensitive to oxidative stress, which is the primary event in any neurological disorder [25]. Peroxidation of membrane lipids may be one of the mechanisms by which ROS contributes to the cascade of events leading to damage of cell membrane and also can induce peroxidation of myelin lipids [26]. When petrosamine is added to the cells, a reduction by approximately 5% in lipid peroxidation is verified in cells submitted to any AlCl3 concentration (Additional file 1: Fig. S4).
2.4 Petrosamine potently inhibits AChE activity after aluminium-induced neurotoxicity in vitroThe increase in AChE activity caused by different concentrations of AlCl3 applied to neuroblastoma cells (Fig. 4) was evaluated with Ellman’s colorimetric assay. AChE activity increased 48.9%, 195.7% (p < 0.05) and 314.6% (p < 0.001) upon treatment with 50, 200 and 1000 µM of AlCl3, respectively, in comparison to life control activity levels. One of the leading causes of the observed decline of the acetylcholine concentration in the brain of AD patients is due to the activity of AChE, which regulates the termination of the synaptic signal by hydrolysing the neurotransmitter acetylcholine secreted in the inter-synaptic cleft in the central nervous system [27]. AlCl3 appears to mimic AD-like neuropathology by altering AChE activity. This parameter has not yet been described in the literature to be affected in the neuroblastoma cell line SH-SY5Y, under the effect of AlCl3, validating the in vitro AD model used in this study. It is nevertheless acknowledged, in the literature, that Al can lead to a higher activity of AChE, decreasing the levels of acetylcholine release [28], which agrees with the results described here. To date, AChE inhibition has been the most widely used therapeutic target for symptomatic improvement in AD, since a cholinergic deficit is commonly observed in this disease [27]. As observed in Fig. 4, petrosamine leads to an intense decline of AChE activity to values lower than in the life control, possibly restoring the levels of acetylcholine neurotransmitter, after neurotoxicity induced in SH-SY5Y cells.
Fig. 4
Fluorescence microscopy of acridine orange (AO)/propidium iodide (PI) assay showing SH-SY5Y cells exposed to different concentrations of AlCl3 for 24 h. a Life Control; b Death Control (30% DMSO); c 50 μM AlCl3; d 50 μM AlCl3 and petrosamine; e 200 μM AlCl3; f 200 μM AlCl3 and petrosamine; g 1000 μM AlCl3; h 1000 μM AlCl3 and petrosamine. Scale bar = 50 μm. Graph i represents the number of viable cells (green), early apoptotic cells (yellow), and late apoptotic/necrotic cells (red) in each condition. * p < 0.05, ** p < 0.01, **** p < 0.0001
2.5 Petrosamine can counteract most negative effects caused by AlCl3 on zebrafish development parametersTo evaluate in vivo the neurotoxicity provoked by AlCl3, several phenotypic parameters of zebrafish embryo development were assessed, including the survival and hatching rate, the yolk volume and the cardiac frequency. Zebrafish embryos were exposed to AlCl3 (50 and 200 µM) until 72 hpf and then treated with petrosamine (0.05 mg/ml) until 120 hpf. The survival rate was evaluated daily during this time. As shown in Fig. 5a, the survival rate of embryos exposed to either concentration of AlCl3 decreased by 20% in the initial 24hpf and addition of petrosamine did not influence this parameter. Zebrafish embryos typically hatch between 48 and 72 hpf [29], so this parameter was only monitored up to 72 hpf indicating the direct effect of exposure to Al. As shown in Fig. 5b, there is a trend of delayed hatching rate of zebrafish embryos exposed to AlCl3, in a dose-dependent manner, when compared to the control. Capriello et al. observed that Al presence led to hatching levels always lower than to the control and, surprisingly, this effect negatively correlated with AlCl3 concentration [30]. The toxicity mechanisms of Al are not sufficiently elucidated and these authors hypothesize that sublethal concentrations of Al differentially affect movement parameters and neuroblast differentiation, with only low levels of metal effecting alterations, thereby allowing extensive sublethal damage, while high levels of metal are capable of activating protective mechanisms.
Fig. 5
Activity of acetylcholinesterase (AChE) enzyme in SH-SY5Y cell line after 24 h of incubation with AlCl3 and further 24 h of petrosamine incubation. * p < 0.05 and *** p < 0.001 when compared to the life control
Epiboly occurs during gastrulation and is the first morphogenetic cell movement during zebrafish embryogenesis [29]. In normal conditions, zebrafish embryo’s epiboly at 5.3 hpf is roughly 50% [31]. The epiboly percentage of control embryos and embryos submitted to 50 and 200 µM of AlCl3 treatment was measured at 6 hpf (Fig. 6). The control was at 75% of epiboly, while zebrafish embryos exposed to 50 and 200 µM concentrations were at 45% (p < 0.05) and 27% (p < 0.001) of epiboly, respectively. This striking difference indicates that the embryos exposed to AlCl3 had a slower development, already verified at 6hpf.
Fig. 6
Analysis of zebrafish embryo development during embryotoxicity assay. a Survival rate after exposure to AlCl3 (50 and 200 μM), with or without treatment with petrosamine (0.05 mg/ml); b hatching rate of zebrafish embryos exposed to AlCl3 (50 and 200 μM)
The zebrafish embryo relies solely on a finite amount of nutrient reserve present on the yolk, which presents an opportunity to gain insight if AlCl3 exposure compromises this phenotypic parameter [32]. Measurement of the yolk dimensions of individual zebrafish embryos over the first 120 hpf can be used to track overall pace of embryo development. At 48 hpf, the yolk volume of the embryos treated with either conditions of AlCl3 was observed to be higher than the control (Additional file 1: Fig. S5). However, only at 120 hpf was identified a statistical significance for embryos submitted to treatment with 50 µM of AlCl3 (p < 0.01). This has not yet been described for Al-mediated toxicity, but Aldavood et al. verified that the yolk sac areas, at 72 hpf were significantly bigger in zebrafish embryos submitted to cadmium exposure as compared to age-matched controls [33]. The effect of AlCl3 on the yolk volume translates into a slower or impaired development of the zebrafish. If the yolk volume does not reduce overtime, the energetic needs during normal development are not being met. We observed that Al toxicity was more marked in larval compared to embryo stages. This probably occurs due to the absence of the chorion in larvae, which in the early stage of development exerts a good protection for the embryo and may somewhat reduce the exposure to the metal. When petrosamine (0.05 mg/ml or 0.01 mM) is added at 72 hpf, it limits this tendency in the yolk volume at 96 and 120 hpf, showing a great impact counteracting AlCl3-mediated toxicity and helping to accelerate the organism development.
The regular zebrafish heart rate ranges 140 to 180 beats per minute (BPM), considerably similar to the human fetal cardiac frequency (130–170 BPM) [34]. The exposure to AlCl3 greatly diminished the cardiac frequency and, as early as 24 hpf, zebrafish embryos exposed to AlCl3 exhibit a cardiac frequency lower than control embryos by approximately 50 BPM (Additional file 1: Fig. S6). There is usually a 20 to 50 BPM increase in heart rate associated with acquired swimming activity as early as day 5, suggesting that neural mechanisms involved in cardiac frequency acceleration are functional in early development [34]. This was not observed in zebrafish larvae exposed to AlCl3, suggesting compromised neural machinery that ultimately affects the cardiac function, through the parasympathetic and sympathetic branches of the autonomic nervous system [35]. The embryos also developed sublethal phenotypic alterations, such as pericardial edema (not shown). Monaco et al. also reported a reduction in cardiac frequency of zebrafish embryos treated with 100 µM of AlCl3 up to 48 hpf, in agreement with our results [19]. Similarly, Gouva et al. submitted zebrafish embryos to 0.05 mg/ml or 0,01 mM.Al treatment and registered a cardiac frequency around 144 BPM [36]. In our study, petrosamine was added at 72 hpf but it did not significantly impact against Al-induced cardiotoxicity in zebrafish embryos (Additional file 1: Fig. S3).
2.6 Petrosamine counteracts apoptosis induced by AlCl3 in zebrafish larvaeControl larvae with a normal phenotype (Fig. 7a) and treated larvae with either 50 µM (Fig. 7b) or 200 µM (Fig. 7d) of AlCl3 were exposed to acridine orange to evaluate apoptosis by fluorescence microscopy. A physiological presence of apoptotic cells can hardly be observed in control larvae. However, a pattern of apoptotic spots (indicated by arrows) was observed in the head of the larvae with 120 hpf exposed to both concentrations of AlCl3. Interestingly, when larvae are treated with petrosamine (0.05 mg/ml or 0.01 mM) after neurotoxicity induced with AlCl3 50 µM (Fig. 7c) and 200 µM (Fig. 7e), no apoptotic spots are observed. In order to perceive which concentration of AlCl3 was causing more apoptosis, the total fluorescence of each green spot was measured using ImageJ software. As it is verified in Fig. 7f, the 50 µM concentration of AlCl3 induces a higher level of physiological apoptotic spots in the larvae’s head, as seen by an extremely higher fluorescence intensity, compared to the control and to the concentration of 200 µM of AlCl3.
Fig. 7
Epiboly of zebrafish embryos exposed to 50 and 200 µM of AlCl3 at 6 hpf. *p < 0.05 and ***p < 0.001 when compared to the control
Apoptosis levels were studied in the zebrafish larvae, with acridine orange, and a mosaic of apoptotic cells in the larvae head was observed. The lateral line is a sensory system used by aquatic animals to detect water flow. This morphological structure is modulated by sensory organs and neuromasts [37]. Neuromasts are innervated by sensory neurons, situated in two ganglia, that leads to the anterior lateral line projected in the head and posterior lateral line along the body. Thus, neuromasts are clusters of superficial hair cells, allowing fish to sense mechanical changes in water, playing a crucial role in behaviours such as rheotaxis, predator avoidance and schooling [37]. The localization of apoptotic cells detected coincides with that of neuromasts, according to the observed pattern of green spots. Additionally, the total fluorescence intensity registered with 50 µM AlCl3 exposure is much stronger than with 200 µM AlCl3, which is similar to that of the control. This indicates that the larvae exposed to 50 µM of AlCl3 have several apoptotic cells in the neuromasts, which suggests that the detection of water movement in these 5-day old larvae may be ineffective. Capriello et al. also reached a similar conclusion, in a study in which they evaluated apoptosis levels in zebrafish embryos treated with AlCl3. The authors verified an increase in apoptosis in the head and tail in the larvae exposed to 50 µM AlCl3, while those exposed to higher doses exhibited a decrease in the number of apoptotic cells [30]. Also, they tested the larvae motility and concluded that motility drastically decreased in embryos exposed to 50 µM AlCl3 but tended to increase in larvae exposed to high concentrations [30]. This is in good agreement with our results, where 50 µM of AlCl3 lead to apoptosis in neuromasts.
2.7 Petrosamine diminishes AChE activity after aluminium-induced neurotoxicity in vivoAnother harmful effect of Al on the nervous system of zebrafish embryos is the alteration of AChE activity, as observed in Fig. 8, also a hallmark in AD. In fish, AChE activity is essential for muscle function and behaviour [38]. This study examined the effects of AlCl3 exposure of AChE activity on whole zebrafish embryos (Fig. 8). Zebrafish embryos submitted to 50 µM of AlCl3 had an increased AChE activity by 67% (p < 0.0001). Senger et al. also verified a significant increase in AChE activity in the brain of adult zebrafish following exposure to AlCl3. The AlCl3 concentrations tested (50–250 μM) increased AChE activity by 28 to 33% [39], which is in line with our results. Senger et al. observed a smaller increase in AChE activity, since they targeted the enzyme only in the brain, excluding the remaining AChE present in the body. Oliveira et al. also reported a dose-dependent increase in enzymatic activity in adult fish Oreochromis niloticus after exposure to AlCl3 and Al2(SO4)3, measured both in individual brain and in individual muscle [40]. However, we did not verify an increase in the activity of this enzyme when the embryos were exposed to 200 µM of AlCl3. This occurred probably to the fact that the Al exposure was performed until 72 hpf, the embryonic stage, where the embryos are protected by the chorion, a biological structure surrounding the embryo until hatching to prevent toxins from entering in contact with the embryo [41]. Therefore, the ions in the highest concentration of AlCl3 may accumulate in the chorion, safeguarding the embryos from Al neurotoxicity. Petrosamine was applied at 72 hpf after Al exposure leading to an AChE activity decrease by 92%, showing high AChE inhibition by this natural compound (Fig. 9).
Fig. 8
Acridine orange (AO) staining in whole body zebrafish larvae with 120 hpf.; a Control larvae; b larvae exposed to 50 μM AlCl3; c larvae exposed to 50 μM AlCl3 and petrosamine; d larvae exposed to 200 μM AlCl3; e larvae exposed to 200 μM AlCl3 and petrosamine; and f Total fluorescence of green spots indicating identified apoptotic cells. Particular fluorescent cellular accumulations are indicated by arrows. Scale bar = 500 μm
Fig. 9
AChE activity of whole 120 hpf embryo, after exposure to two different concentrations of AlCl3 (50 µM and 200 µM) during 72 hpf and then treated with petrosamine (0.05 mg/ml) until 120 hpf. Statistical significance as compared to life control, ****p < 0.0001
Lin et al. tested the AChE inhibition of fucoxanthin, a natural carotenoid abundant in edible brown seaweeds, fucoxanthin significantly reversed the scopolamine-induced increase of AChE activity in the hippocampus and cortex of mice [42]. Additionally, Pam et al. tested derivatives of fascaplysin, an alkaloid isolated from a marine sponge Fascaplysinopsis, suggesting that one of the derivatives tested could penetrate the blood–brain barrier and be retained in the central nervous system and inhibit AChE with an IC50 value of 1 μM. Thus, Pam et al. verified that the fascaplysin derivative prevented cholinergic dysfunction and, consequently, prevented cognitive impairments by inhibiting AChE activity., in two AD mice models [43]. Nie et al. tested a marine fungal metabolite named butyrolactone I in vivo by implementing a cognitive deficit model in adult zebrafish, evoked by intraperitoneal injection of AlCl3, The authors verified that the AlCl3 treatment increased AChE activity, while butyrolactone I dose-dependently ameliorated AlCl3-induced cognitive deficits in zebrafish [
留言 (0)