
記住我
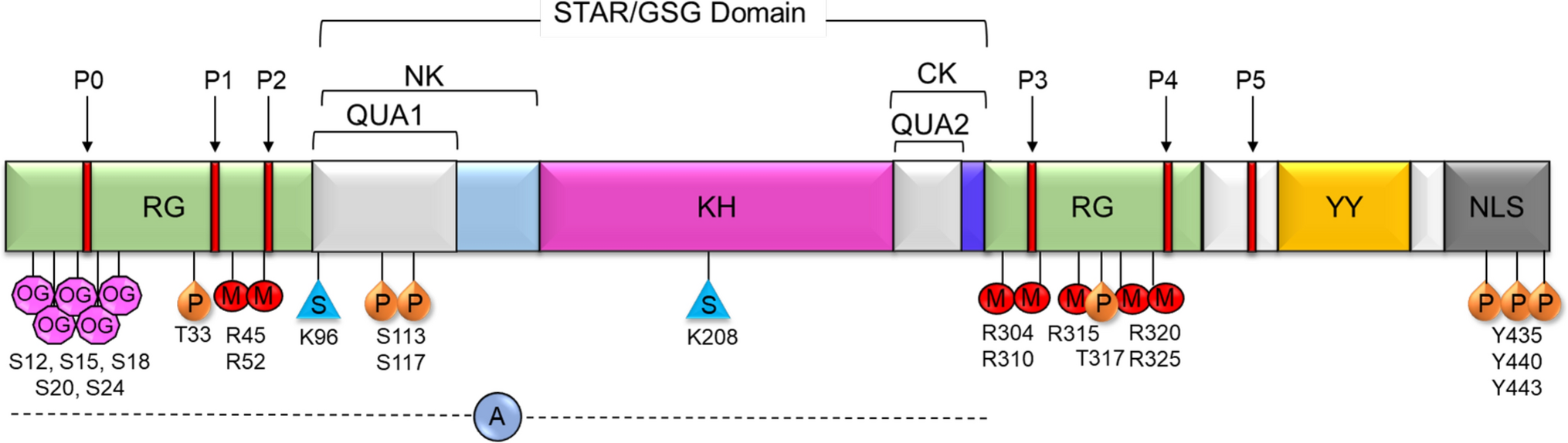
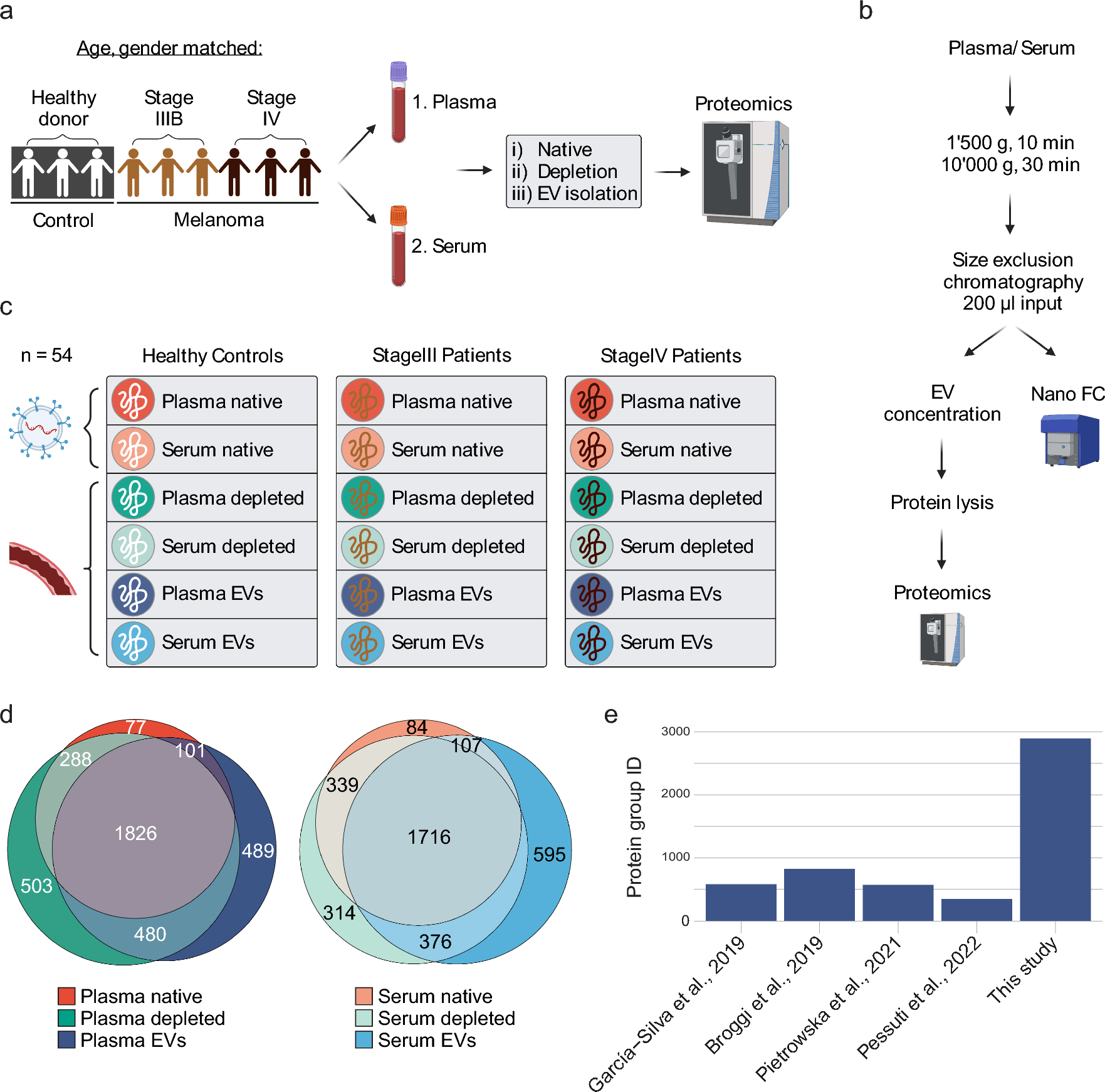
Src-Associated in Mitosis 68 kDa Protein (Sam68), also known as KH Domain-Containing, RNA-Binding, Signal Transduction-Associated Protein 1 (KHDRBS1), belongs to the signal transduction and activation of RNA (STAR) family of proteins [1]. Sam68 plays key roles in cell proliferation, differentiation, development, signal transduction, and various aspects of RNA metabolism such as transcription, alternative splicing, and nuclear export [2, 3]. Sam68 contains a heteronuclear ribonucleoprotein particle K (hnRNP K) homology (KH) domain like other RNA-binding proteins in the STAR family. The KH domain is critical for the recognition and subsequent binding of Sam68 to the 3′ U(C/A)A(C/U) moiety commonly found on RNA. The KH domain is flanked by the highly conserved QUA1 region within the N-terminus (NK) and QUA2 region within the C-terminus (CK) of KH domain [3] (Fig. 1). The QUA1 region plays an important role in Sam68 dimerization, while the QUA2 region is responsible for recognizing a specific RNA sequence and stabilizing the dimer orientation via contact with the QUA1 and KH regions [3]. Together, the NK/QUA1, KH, and CK/QUA2 regions make up the STAR domain or GRP33/SAM68/GLD-1 (GSG) domain that is essential for RNA binding and homodimerization [1]. In addition to the STAR/GSG domain, Sam68 contains various regions that mediate protein–protein interactions as well as multiple posttranslational modification sites (Table 1). The C-terminus of Sam68 contains tyrosine residues which are phosphorylated, which mediate binding to SH2 domain-containing proteins (Table 1) [2]. The protein’s six proline-rich regions allow for binding to SH3 domains of Src kinases, which regulate a wide variety of cellular functions such as cell proliferation, survival, and migration [4,5,6]. The tyrosine phosphorylation-dependent SH2 domain protein binding and proline-rich region-dependent SH3 domain binding have been shown to inhibit Sam68’s interaction with RNA and facilitate its function as an adaptor molecule in signaling involved in cell cycle progression and cell proliferation (Table 1) [2]. In addition to phosphorylation, other posttranslational modifications, including O-GlcNAcylation [7], arginine methylation [8], SUMOylation [9], and acetylation [10], control cellular functions of Sam68 (Table 1). Lin et al., reported that the O-GlcNAcylation of Sam68 plays a role in modulating lung cancer cell migration and invasion [7]. Arginine methylation of Sam68 by the protein arginine methyltransferase 1 (PRMT1) has been shown to decrease the RNA-binding ability of Sam68, modulating its protein interactions and controlling its localization and function [8, 11]. Sam68 is SUMOylated by SUMO ligase PIAS1, which results in the repression of cyclin D1 leading to G1 arrest and inhibition of cell proliferation (Table 1). However, it is unclear whether SUMOylation of Sam68 has any effect on the RNA-binding affinity of Sam68 and protein–protein interactions [9]. The RNA-binding domain is required for Sam68 apoptosis induction, while acetylation of Sam68 enhances its RNA-binding ability and may upregulate pro-apoptotic proteins [9]. Sam68 also contains a nuclear localization signal (NLS) in its C-terminus which allows the protein to move between the cytoplasm and nucleus of the cell, as well as execute its many transcriptional and posttranscriptional regulatory roles [2].
Fig. 1
Schematic representation of Sam68 domains and posttranslational modifications. P0–P5—proline-rich regions, RG—arginine/glycine-rich regions, QUA1 and QUA2—domains flanking KH domain where QUA1 mediates dimerization of Sam68, KH—hnRNP K homology domain, NK—N terminal of KH, CK—C terminal of KH, YY—tyrosine phosphorylation sites, NLS—nuclear localization signal. Known posttranslational modifications of Sam68 are annotated at the respective residues. OG—O-GlcNAcylation, P—phosphorylation, M—methylation, S—SUMOylation, A—acetylation region
Table 1 Posttranslational modifications of Sam68 and their biological significanceThe lion’s share of the studies on Sam68 focuses on its roles in RNA binding and cancer [1, 12, 13]. Sam68 has also been shown to play a critical role as an adaptor protein in various signaling pathways such as insulin [14, 15], leptin [14, 15], T-cell receptor (TCR) and B-cell receptor (BCR) [14, 16], tumor necrosis factor (TNF) [6], and toll-like receptor (TLR) [17]. Recent findings suggest that higher expression levels of Sam68 are not only associated with various human cancers [1, 2, 18], but also with inflammatory diseases such as inflammatory bowel disease [19], arthritis [20], and arterial injury [21]. For example, the high levels of Sam68 observed in patients with ulcerative colitis (UC) correlated with higher levels of apoptosis and NF-κB activation [19]. As such, Sam68’s role in driving inflammation through NF-κB signaling and inhibition of wound repair processes has been linked to worsened patient outcomes across various inflammatory diseases [19, 21]. Understanding Sam68-dependent inflammatory signaling pathways and delineating the multifaceted effects of Sam68 on inflammation may improve patient prognosis and drive therapeutic development for inflammatory diseases. In this review, we highlight Sam68’s adaptor role in inflammatory diseases and discuss its potential as a therapeutic target.
Sam68 as an adaptor proteinAdaptor proteins are essential for the propagation, amplification, and regulation of cellular signals [22]. In inflammatory signaling, adaptor proteins may play an essential role in both the specificity and diversity of cellular responses to inflammation, as well as to resolve inflammatory signaling before it becomes pathologic [23]. Sam68, first identified as an RNA-binding protein, has been shown to serve as an adaptor protein in an array of inflammatory signaling events, notably at the upstream-receptor level.
Role of Sam68 in TCR signalingThe TCR enables T cells to recognize foreign antigens and signal to mount immune and inflammatory responses. TCR activation initiates a signaling cascade which triggers the recruitment of several cytoplasmic proteins to the receptor [24, 25]. The presence of proline-rich regions allows for Sam68’s interactions with SH3 domain containing adaptors such as Grb2 (Fig. 2A) [26] and the Src family tyrosine kinases, Fyn and Lck [27, 28]. These interactions may occur at the basal state, as they do not involve any stimulus-dependent posttranslational modifications. The six proline-rich domains throughout Sam68 are notably not clustered to one particular terminus, resulting in a spatially heterogeneous distribution of SH3 domain-binding motifs on Sam68 [29]. It is also possible that the conformational changes in the three-dimensional structure of Sam68, upon carboxy-terminal SH2 binding, may allow for a differential display of one or more proline-rich regions. Importantly, this would promote more efficient binding to SH3 domain containing proteins or binding to more than one SH3 domain containing protein, facilitating an essential adaptor/scaffolding role in propagating downstream TCR signaling. Furthermore, the binding of one SH3 domain-containing protein may also expose additional proline-rich regions or result in tyrosine phosphorylation of Sam68, thus allowing additional recruitment of both SH3 and SH2 domain containing proteins.
Fig. 2
A Schematic representation of the role of Sam68 in TCR signaling. Upon TCR activation, multiple tyrosine residues are phosphorylated at immunoreceptor tyrosine-based activation motifs (ITAM) of the TCR–CD3 complex by the Src-family protein-tyrosine kinase LCK. Both LCK and Fyn contain SH3 and SH2 domains which may bind to naïve and phosphorylated Sam68, respectively. This is followed by the recruitment and activation of ZAP-70 and the scaffolding protein, SLP-76, which may bind to phosphorylated Sam68 through their SH2 domains. Signaling through SLP-76 and Grb2, which contain both SH2 and SH3 domains to facilitate Sam68 binding, will lead to activation of the MAP kinase pathway. The engagement of CD28 and co-stimulation of the TCR will lead to the activation of PI3 kinase and PKC, resulting in MAP kinase and IKK–NF-κB pathway activation. Activated IKK1 has also been shown to phosphorylate Sam68 in the nucleus and promote its binding to NF-κB p65 and activate CD25 expression. B Schematic representation of the role of Sam68 in BCR signaling. Upon BCR activation, through antigen binding, kinases such as Lyn and Btk, which contain both SH3 and SH2 domains, are recruited to the receptor complex. Sam68 may bind to the receptor complex through a proline-rich region—SH3 interaction in its naïve state, and when phosphorylated, Sam68 may bind to other proteins such as Syk through SH2 domain interactions. Downstream signaling proteins such as Grb2, GAP, and PLCγ contain both SH2 and SH3 domains which may facilitate binding to Sam68 and lead to Ras activation. Phosphorylated Sam68 will also interact with the PI3K’s SH2 domain to control downstream signaling events in B cell receptor signaling
In contrast, Sam68’s binding to SH2 domain containing proteins is dependent on its tyrosine phosphorylation, which may happen only after TCR activation. TCR engagement results in the recruitment of several proteins along with multiple kinases such as Lck and Fyn, both of which contain SH3 and SH2 domains to facilitate the binding of non-phosphorylated and phosphorylated forms of Sam68. It is possible that the initial binding of Sam68 to the TCR complex is mediated through its interaction with the SH3 domain-containing proteins followed by its tyrosine phosphorylation. It is observed in in vitro studies that once tyrosine phosphorylation has occurred, Sam68 may also bind to the adaptor proteins such as ZAP-70 and SLP-76 and phosphoinositide 3-kinase (PI3K) as well as activate mitogen-activated protein kinase (MAPK) pathways (Fig. 2A) [25, 30]. Thus, through the interactions via proline-rich regions and tyrosine phosphorylated regions, Sam68 may perform its adaptor function in linking multiple proteins’ binding to the ζ chain of the TCR’s CD3, as well as to the intracellular domains of CD4/CD8, CD28, and linker for activation of T cells (LAT) (Fig. 2A).
Absence of Sam68 has been shown to significantly reduce the TCR-induced inflammatory interferon (IFN)-γ production in T helper 1 (Th1) cells. This was primarily attributed to Sam68’s ability to bind and inhibit microRNA-29, which inhibits the transcription factors T-bet and Eomes which control IFN-γ expression in T cells [31]. It remains to be investigated whether the adaptor function of Sam68 at the TCR also contributes to IFN-γ expression in T cells. Furthermore, Sam68 has been shown to be an essential non-Rel component of the NF-κB complex inducing CD25 expression in T cells [16] (Fig. 2A). T-cell activation results in IKKα-dependent serine phosphorylation of nuclear Sam68 and its binding to the NF-κB p65 subunit, and subsequent binding of this Sam68-p65 complex to the CD25 promoter. This alludes to an important nuclear-adaptor role of Sam68 in regulating gene expression. Because Sam68-p65 binding does not directly involve SH2/SH3 domain-dependent interactions, knowledge of the precise mechanism of Sam68-p65 binding in T cells is expected to reveal novel binding mechanisms through which Sam68 executes its adaptor role in the nucleus. Notably, some contradictions exist within the cytoplasmic adaptor role of Sam68 in TCR signaling. In a well-performed study, Fu et al. [16] showed that siRNA suppression of Sam68 in Jurkat cells did not prevent TCR activation and that PMA/ionomycin stimulation induced IκBα degradation. This raised the possibility that Sam68 may not be essential in cytoplasmic signaling events in T cells leading to NF-κB activation. However, the siRNA-mediated suppression of Sam68 in this study leaves a significant amount of residual Sam68 in the cells, which may suffice for signal transduction. Hence, to derive firm conclusions on the adaptor role of Sam68 in TCR signaling, it is necessary to conduct comprehensive studies using complete Sam68 knockout T cells and delineate downstream signaling following both CD3 activation alone and with CD3 and CD28 co-stimulation. Moreover, given the presence of multiple potential binding partners for Sam68 in the TCR complex and in the nucleus, a clearer understanding of the direct and indirect binding partners of Sam68 is necessary to elucidate the biochemical mechanisms by which Sam68 adapts TCR signaling.
Role of Sam68 in BCR signalingB cells are another major component of the adaptive immune system with a major role in antibody production [32]. They also partake in antigen presentation and secrete multiple cytokines that control immune response [33]. The BCR recognizes and binds to foreign antigens, leading to B cell activation and propagation of signaling via phosphorylation of multiple kinases including Lyn, Btk and PI3K at the BCR complex [34]. Sam68 interacts with the PI3K–SH2 domain through its phosphotyrosine region [34, 35] and the Btk–SH3 domain through its proline-rich regions (Fig. 2B) [36]. The proline-rich regions were also shown to control B cell signal transduction regulating cell growth and death [4]. Lyn kinase has been shown to phosphorylate Sam68 [37], but its direct interaction with Sam68 in B cells remains to be demonstrated. Lyn kinase has also been shown to be required for the activation of Syk kinase, which involves trans- and auto-phosphorylation events [38]. It is highly likely that the activated Syk kinase may interact with and phosphorylate Sam68 propagating downstream BCR signaling involving Lyn and Btk kinases [36, 38,39,40] based on the potential of Syk to phosphorylate Sam68 in vitro [41]. Sam68 has also been shown to interact with several other SH2 and SH3 domain-containing proteins that are associated in the BCR complex such as Grb2, PLC-gamma, and RAS GTPase activating protein (Ras-GAP) [42, 43]. Through this multiprotein binding ability, Sam68 is likely to control downstream signaling pathways activated following BCR stimulation such as NF-κB, nuclear factor of activated T-cells (NFAT), p38, JNK, ERK, and AKT. The role of Sam68 in adapting these kinases/pathways in BCR signaling remains a vast area inviting comprehensive investigation.
Role of Sam68 in insulin receptor (IR) signalingInsulin binding to the IR results in tyrosine phosphorylation of insulin receptor substrate 1 (IRS1) and downstream signaling cascade activation [44]. Sam68 has been shown to bind to IRS1 at the basal state and this interaction enhances following insulin stimulation, with an associated increase in Sam68 protein level both in the cytoplasm and at the IR. At the IR, Sam68 associates with PI3K via the SH2 domain of the p85 subunit (Fig. 3), a main downstream constituent of insulin signaling [46]. Insulin signaling also causes tyrosine phosphorylation of Sam68 enabling its interaction with SH2 domain containing proteins such as GAP and PI3K, both in vitro and in vivo [26, 35, 45]. Sam68 also has the ability to bind and recruit p120GAP to the PI3K pathway [26], possibly through interaction with the SH2 domain. The simultaneous association of Sam68 with p85 PI3K and GAP help to bring these two molecules together, thus linking PI3K with the RAS pathway in IR signaling (Fig. 3) [35, 47]. On the other hand, Sam68 is known to be associated with the SH3 domains of Grb2 independent of insulin stimulation in vivo; however, insulin stimulation has been shown to promote Sam68 phosphorylation and its association with the SH2 domains of GAP. This in turn promotes association of GAP with the Grb2–SOS complex. Both of these complexes lead to the downstream activation of the MAP kinase pathway, influencing metabolic and mitogenic gene expression (Fig. 3) [26]. Overall, current evidences show that Sam68 is a substrate phosphorylated in insulin signaling, and that it also acts as an adaptor linking multiple proteins at the IR. Most of the current knowledge on the role of Sam68 in IR signaling are derived from in vitro studies using cell lines and their physiological significance remains to be validated through in vivo studies. Possible IR-independent adaptor roles of Sam68 in the cytoplasm that may control inflammatory signaling and insulin resistance, as well the potential nuclear roles of Sam68 regulating insulin-dependent gene expression, are interesting areas pending investigation. The relationship between inflammation and insulin resistance has been well studied [48,49,50], but there remain further avenues of investigation. For example, pro-inflammatory cytokines are also known to induce the activation of serine and threonine kinases, which can in turn cause inhibitory phosphorylation of IRS1 and impede downstream signaling by IRS1 [
留言 (0)