
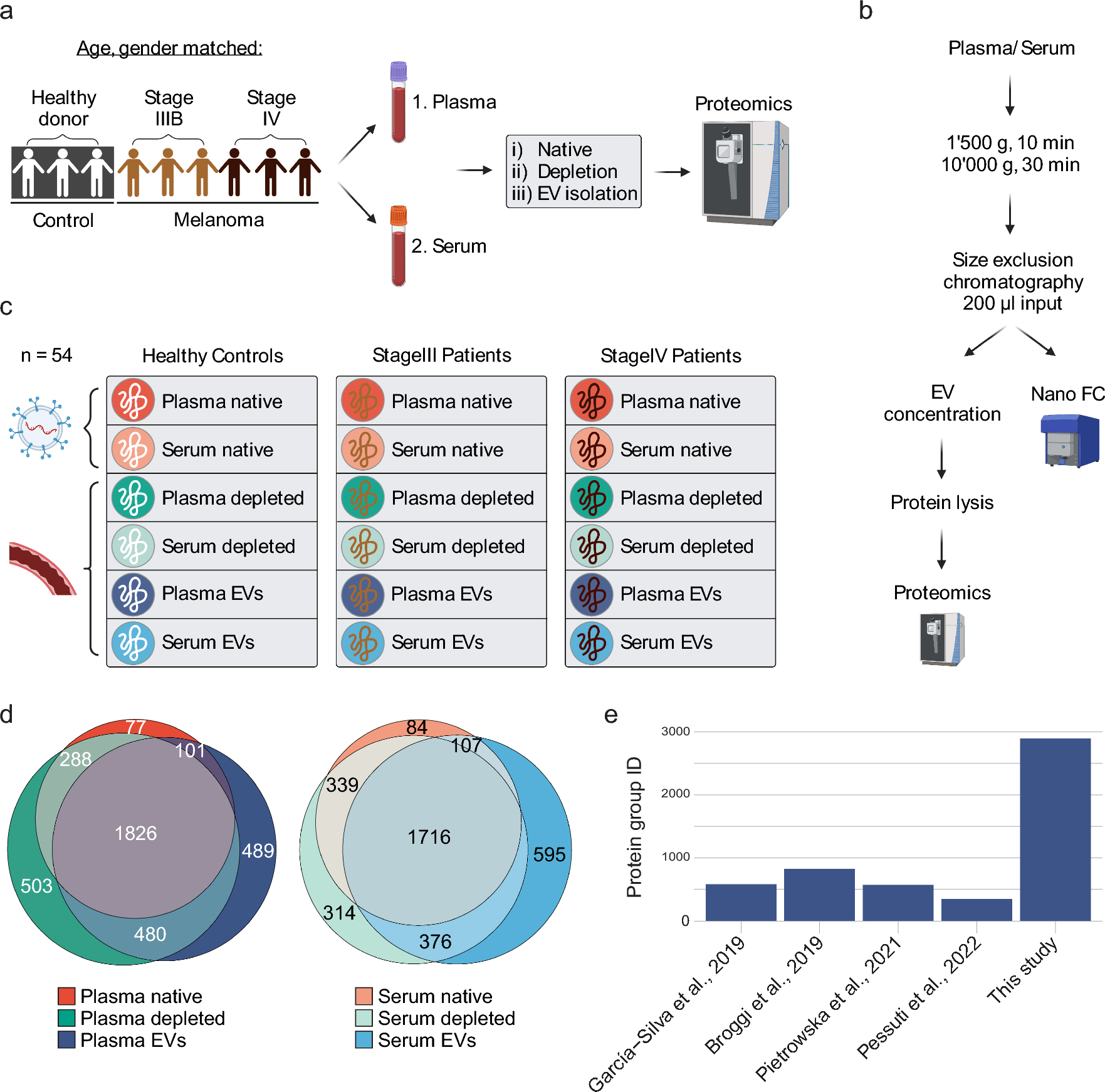
記住我
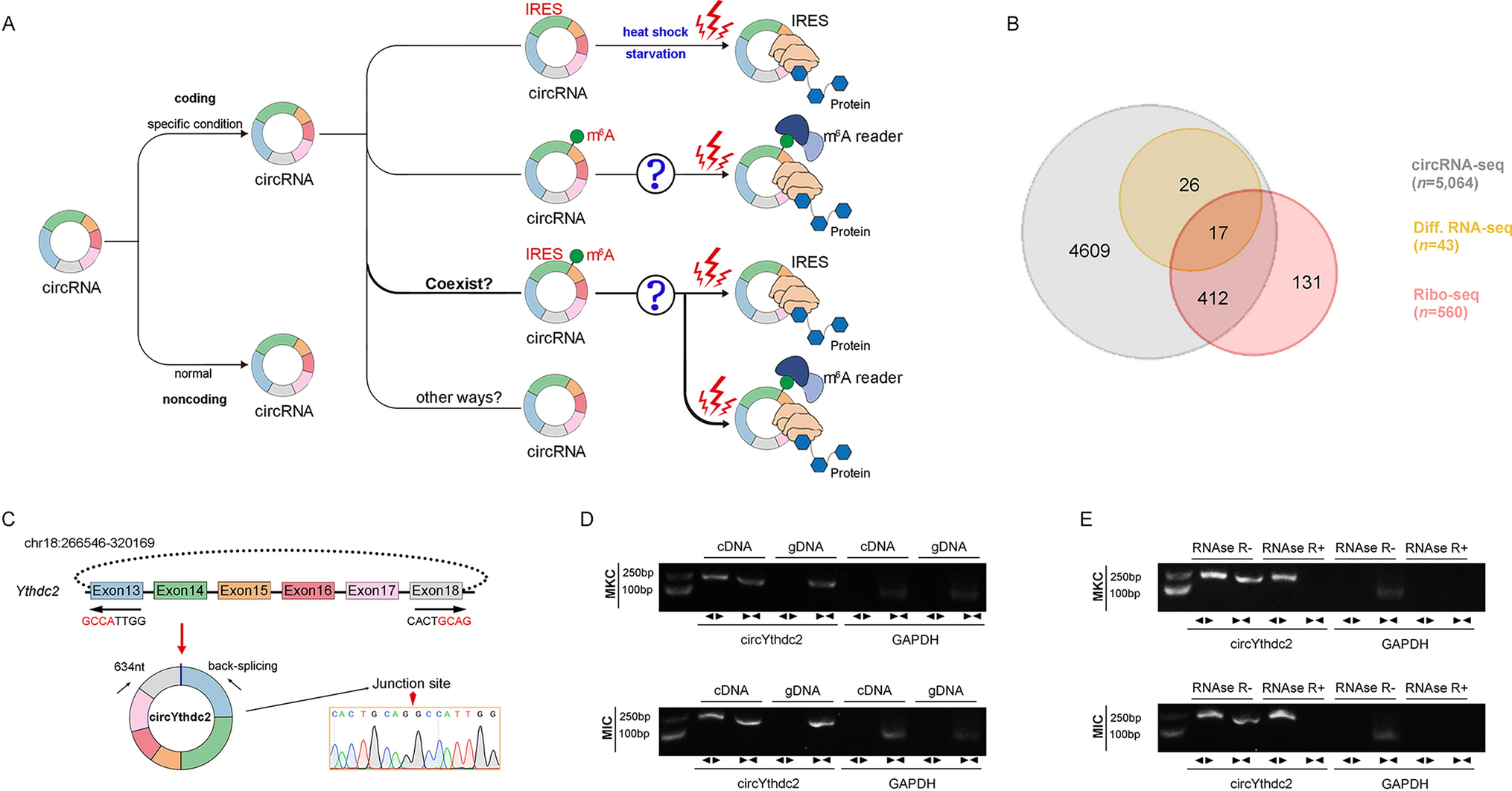
The above-mentioned question about whether circRNA translation is widespread, its translation mode and activation ways are still unknown to a large extent (Fig. 1A), and we are very interested in these currently unknown questions, so we have studied the above-mentioned problems by using the representative fish of lower vertebrates as a model. First, we treated miiuy croaker with SCRV virus, then performed RNA-seq analysis to compare the circRNA expression levels between SCRV-treated and untreated spleen samples (GenBank accession no. PRJNA685924). In total, 5,064 distinct circRNA candidates were found. At the same time, we also carried out Ribo-Seq and obtained 560 candidate circRNAs. By crossing with the differential groups obtained from the above SCRV infection, we obtained some candidate circRNAs (Fig. 1B). Among them, we found circular Ythdc2 (circYthdc2) RNA was most differentially expressed and had coding potential.
Fig. 1
Expression profiles and characterization of circYthdc2. A Schematic diagram of circRNA translation ways and its selection conditions. B Strategies used for circRNA-seq and ribosome profiling (Ribo-seq). The gray strips represented the total circRNAs by circRNA-Seq. The orange strips represented the differential circRNAs upon SCRV treatment. The red strips represented the circRNAs with potential translation ability by Ribo-seq. C We confirmed the head-to-tail splicing of circYthdc2 in the circYthdc2 RT-PCR product by Sanger sequencing. D RT-PCR validated the existence of circYthdc2 in MKC and MIC cell lines. CircYthdc2 was amplified by divergent primers in cDNA but not gDNA. GAPDH was used as a negative control. E The expression of circYthdc2 and linear Ythdc2 mRNA in both MKC and MIC cell lines was detected by RT-PCR assay followed by nucleic acid electrophoresis or qPCR assay in the presence or absence of RNase R. All data represented the three independent triplicated experiments
The transcriptome sequencing revealed that circYthdc2 was 634nt in length (Fig. 1C). Using miiuy croaker whole genome library to perform blast analysis on the host gene Ythdc2 [28, 29], it was found that the Ythdc2 gene was located on chromosome 18, which was composed of 30 exons, and circYthdc2 was self-cyclization from exon13 to exon18. To confirm the objective existence of circYthdc2, first, circYthdc2 divergent primers were designed for RT-PCR amplification, and the amplified products were subjected to Sanger sequencing to confirm that circYthdc2 was spliced from the head to the tail (Fig. 1C). Then, we used convergent primers to amplify Ythdc2 gene and divergent primers to amplify circYthdc2. cDNA and gDNA were extracted separately from MKC and M. miiuy intestine cells (MIC) and subjected to RT-PCR and agarose gel electrophoresis assays. The results shown in Fig. 1D indicated that circYthdc2 was amplified from cDNA using only divergent primers, whereas no amplification product was observed from gDNA. Considering that stability was a crucial characteristic of circRNAs, we thus employed RNase R to confirm the stability of circYthdc2. The results from the agarose gel electrophoresis assay showed that circYthdc2, rather than linear Ythdc2 or GAPDH, resisted digestion by RNase R (Fig. 1E). Among the aforementioned cell lines, MKC and MIC cells showed the lowest and highest expression of circYthdc2, respectively. To confirm the reliability of this result, we conducted in vivo and in vitro experiments to detect the changes in the expression level of circYthdc2 under SCRV infection. At the same time, we also detected the expression of the host gene of this circRNA, Ythdc2, after SCRV infection. The results of qPCR experiments showed that Ythdc2 was significantly up-regulated in miiuy croaker spleen tissue treated with SCRV and Poly (I: C) at different time points (Fig. S1A). The level of circYthdc2 significantly increases during the initial period of SCRV infection or Poly (I: C) stimulation, but subsequently undergoes a significant decrease (Fig. S1A). In addition, SCRV-treated M. miiuy kidney cells (MKC) further confirmed the significant expression of circYthdc2 and Ythdc2 (Fig. S1B). We then evaluated the expression levels of circYthdc2 in MKC, M. miiuy spinal marrow cells (MSpC), MIC, M. miiuy brain cells (MBrC), M. miiuy muscle cells (MMC), and M. miiuy liver cells (MLC), and these cell line samples were divided into two groups, one treated with RNase R and the other untreated. The mRNA expression of Ythdc2 decreased significantly after RNase R treatment in all aforementioned cell lines, while the expression of circYthdc2 almost did not decrease in all aforementioned cell lines (Fig. S1C). Therefore, we selected MKC and MIC to investigate the function and regulatory mechanism of circYthdc2. Moreover, the half-time of the circYthdc2 transcript was significantly longer than Ythdc2 mRNA after being treated with actinomycin D, which suppressed RNA transcription (Fig. S1D).
In addition, we detected the distribution of circYthdc2 by cytoplasmic nuclear fractionation experiments and found that circYthdc2 was primarily localized in the cytoplasm (Supplementary Fig. 1E). Accordingly, these results suggested that circYthdc2 was a stable circRNA expressed and primarily distributed in the cytoplasm.
circYthdc2 encodes a 170 amino acid (aa) novel protein, Ythdc2-170aacircYthdc2 has a predicted ORF, which may encode a 170 aa protein using an overlapping start-stop codon “UAG……AUG” (Fig. 2A). Through prediction, we found that IRES sequence exists on circYthdc2. Therefore, to confirm this, we carried out a related dual-luciferase assay and GFP protein activity assay to detect IRES activity, and the experimental results show that the complete IRES sequence predicted from circYthdc2 does have the activity of mediating translation. Once the IRES sequence is deleted, it will lead to a significant decrease in activity (Fig. 2B and C). We constructed the Linear-FL-Ythdc2-AG overexpression plasmid, FLAG-circYthdc2 overexpression plasmid, and a linearized Ythdc2-170aa overexpression plasmid, and the construction pattern of these plasmids is shown in Fig. 2D. To confirm that the 170 aa predicted protein was encoded by circYthdc2, we used the Linear-FL-Ythdc2-AG overexpression plasmid, FLAG-circYthdc2 overexpression plasmid, and a linearized Ythdc2-170aa overexpression plasmid. We used an antibody against the Ythdc2-170aa, which should recognize Ythdc2-170aa. In MKC cells, which have lower endogenous circYthdc2 levels, the transfection circYthdc2 and the linearized Ythdc2-170aa plasmid both resulted in the predicted Ythdc2-170aa band, while the overexpression Linear-FL-Ythdc2-AG plasmid did not. In addition, the Ythdc2-170aa polypeptide cannot be detected under normal conditions, but we found that the Ythdc2-170aa polypeptide was generated in large quantities under the condition of SCRV virus infection (Fig. 2E). These results prove that Ythdc2-170aa encoded this 170aa novel protein, which we termed Ythdc2-170aa. Immunofluorescence using an anti-flag antibody confirmed the cytoplasmic localization of Linear-FLAG-Ythdc2-170aa in MKC cells, as shown in Fig. 2F.
Fig. 2
CircYthdc2 encodes a 170 amino acid (aa) novel protein, Ythdc2-170aa. A Upper panel, the putative ORF in circYthdc2. Lower panel, the sequences of the putative ORF are shown. B The putative IRES activity in circYthdc2 was tested. C Left panel: Full-length or truncated circYthdc2 IRES sequences were cloned before GFP as indicated to construct reporter plasmids. Right panel: The empty vector, and full-length or truncated IRES vector were cotransfected with si-eif4E into HEK293 cells, and GFP signals were detected. D Schematic diagram of FLAG-circYthdc2, Linear-FL-Ythdc2-AG, and Linear-FLAG-Ythdc2-170aa plasmid construction. E Upper panel: The putative Ythdc2-170aa amino acid sequences and antibody generation region were shown as indicated to produce the Ythdc2-170aa antibody. The red amino acids were distinctly formed by the circYthdc2 junction. Lower: FLAG tag antibody was used to detect Ythdc2-170aa expression in MKC cells transfected with the vectors mentioned in Fig. 2D. In addition, Ythdc2-170aa antibody was used to detect Ythdc2-170aa expression in MKC cells after SCRV infection F FLAG-circYthdc2, Linear-FL-Ythdc2-AG, and Linear-FLAG-Ythdc2-170aa plasmids were transfected into MKC cells. Immunofluorescence staining using anti-Flag was performed to show the Ythdc2-170aa cellular localization. Original magnification is 630; all data represent the means ± SE from three independent triplicate experiments. *, p < 0.05; **, p < 0.01
CircYthdc2 and Ythdc2-170aa inhibit host antiviral innate immunityWe designed the two small interfering RNAs (siRNA) against circYthdc2, and the overexpression plasmid of circYthdc2 was constructed to explore the biological function of circYthdc2 (Fig. 3A and B). Consequently, two siRNAs (si-circYthdc2-1 and si-circYthdc2-2) decreased the circYthdc2 expression level, but such siRNAs did not affect the expression level of linear Ythdc2 mRNA in MKC. As si-circYthdc2-1 could induce higher inhibitory efficiency; thus, we selected si-circYthdc2-1 (termed si-circYthdc2) for the subsequent experiment (left panel of Fig. 3C). Moreover, the circYthdc2 overexpression plasmid was successfully constructed, as it significantly increased the circYthdc2 expression levels rather than linear Ythdc2 mRNA in MKC cells (right panel of Fig. 3C). Considering that IFN and ISGs are important antiviral effectors, we focused on investigating the role of circYthdc2 and Ythdc2-170aa in regulating the expression of IFN, ISGs, and inflammatory cytokines. As shown in Fig. 3D, the silence of circYthdc2 increased the expression levels of these genes under SCRV treatment. By contrast, the overexpression of circYthdc2 or Linear-FLAG-Ythdc2-170aa both could significantly inhibit the expression levels of interferon IFN1, inflammatory cytokines (TNF-α), and antiviral genes such as myxovirus resistance protein 1 (Mx1), ISG15, and Viperin after SCRV infection (Fig. 3E).
Fig. 3
CircYthdc2 and Ythdc2-170aa inhibit host antiviral innate immunity. A and B The schematic diagram of siRNAs (A) and circYthdc2 overexpression plasmid structure (B). C qPCR analysis of circYthdc2 and linear Ythdc2 mRNA in MIC cells treated with siRNAs. qPCR analysis of circYthdc2 and linear Ythdc2 mRNA in MKC cells overexpressing circYthdc2. D and E qPCR assays were performed to determine the expression levels of IFN1, TNF-α, Mx1, ISG15, and Viperin in MIC cells transfected with NC or si-circYthdc2 (D) and transfected in MKC cells with circYthdc2 or pLC5-circ and Linear-FLAG-Ythdc2-170aa or pcDNA3.1-FLAG after SCRV infection (E). F circYthdc2 and Ythdc2-170aa promote SCRV replication. MIC cells transfected with NC or si-circYthdc2 and MKC cells were transfected with pLC5-circ or circYthdc2 and pcDNA3.1-FLAG or Linear-FLAG-Ythdc2-170aa for 24 h, respectively, then infected with SCRV at 24 h. The qPCR analysis was conducted for intracellular and supernatant SCRV RNA expression. G MKC cells seeded in 48-well plates overnight were treated with cultural supernatants at the dose indicated for 48 h. Then, cell monolayers were fixed with 4% paraformaldehyde and stained with 1% crystal violet. MKC cells were transfected with Linear-FLAG-Ythdc2-170aa or pcDNA3.1-FLAG. H Effect of circYthdc2 on cell viability after SCRV infection. MIC cells were transfected with NC or si-circYthdc2 for 24 h and then treated with SCRV for 24 h. Cell viability assay was measured. I Ythdc2-170aa counteracts the negative effect of STING. Relative luciferase activities were detected in MKC after cotransfection with STING, MAVS, TRIF, and TBK1 expression plasmid, pRL-TK Renilla luciferase plasmid, luciferase reporters, pcDNA3.1-FLAG or Linear-FLAG-Ythdc2-170aa. J Relative protein levels of STING in MIC cells after transfected with NC or si-circYthdc2 and in MKC cells with pLC5-circ or circYthdc2 and pcDNA3.1-FLAG or Linear-FLAG-Ythdc2-170aa or Linear-FL-Ythdc2-AG. All data represented the mean ± SE from three independent triplicated experiments. *, p < 0.05; **, p < 0.01
Furthermore, we examined the effect of circYthdc2 and Ythdc2-170aa on SCRV replication to explore the biological significance of circYthdc2 and Ythdc2-170aa in SCRV-induced host cells. Detecting SCRV RNA level by qPCR, we found that silence of circYthdc2 significantly inhibited SCRV replication after 24 h of SCRV infection (left panel of Fig. 3F). In addition, both circYthdc2 and Linear-FLAG-Ythdc2-170aa overexpression significantly promoted SCRV replication after 24 h of SCRV infection (right panel of Fig. 3F). In addition, SCRV replication was also monitored by the appearance of CPE (cytopathic effect) in MKC cells. As shown in Fig. 3G, overexpression of Linear-FLAG-Ythdc2-170aa presented more CPE in the cells. These results demonstrated that Ythdc2-170aa can promote SCRV replication. When we investigated the effect of circYthdc2 on the cell viability of MIC cells, we found that the silence of circYthdc2 significantly increased cell viability compared with the control group after 24 h of SCRV infection (Fig. 3H). We want to know which gene in the antiviral signal pathway of teleost is affected by Ythdc2-170aa, so we tested STING, MAVS, TRIF, and TBK1 through the double luciferase reporter gene system. Then, the STING, MAVS, TRIF, and TBK1 plasmid with only CDS; Ythdc2-170aa plasmid; and various reporter gene plasmids were cotransfected into EPC cells. The results showed that Ythdc2-170aa inhibited the activity of reporter genes such as IFN1, NF-κB, IRF3, and ISRE commonly by affecting the activity of STING (Fig. 3I). In order to further verify the correctness of the results of double luciferase experiment, we detected the effect of circYthdc2 on STING protein by Western blotting in MKC and MIC cells. The result shows that silence of circYthdc2 significantly increased the protein levels of STING, and overexpression circYthdc2 and Ythdc2-170aa both significantly decreased the protein levels of STING, and overexpression Linear-FL-Ythdc2-AG did not have any effect to STING protein (Fig. 3J). Collectively, circYthdc2 could inhibit STING-mediated antiviral immune response by promoting the degradation of STING protein, and all the experimental results show that the inhibitory effect of circYthdc2 on antiviral immune response mainly comes from the role of its encoded polypeptide Ythdc-170aa.
Ythdc2 inhibits host antiviral innate immunityThrough prediction, we know that Ythdc2-170aa has only one HELICc domain, and Ythdc2, the host gene of circYthdc2, also has this domain, and their amino acid sequences are consistent. Therefore, we suspect that they may have the same function. We designed the siRNA against Ythdc2, and the overexpression plasmid of Ythdc2 was constructed to explore the biological function of Ythdc2. Consequently, the siRNA (si-Ythdc2) decreased the Ythdc2 protein level and mRNA level in MKC cells (Fig. 4A). We detected whether Ythdc2 could affect the expression level of STING protein by Western blotting. As we predicted, the silence of Ythdc2 significantly increased the protein levels of STING, and overexpression of Ythdc2 significantly decreased the protein levels of STING (Fig. 4B). We also focused on investigating the role of Ythdc2 in regulating the expression of IFN, ISGs, and inflammatory cytokines. As shown in Fig. 4C, the silence of Ythdc2 increased the expression levels of these genes under SCRV treatment. By contrast, the overexpression of Ythdc2 could significantly inhibit the expression levels of interferon IFN1, TNF-α, Mx1, ISG15, and Viperin after SCRV infection (Fig. 4D). Then, the STING plasmid with only CDS, Ythdc2 plasmid, and various reporter gene plasmids were cotransfected into EPC cells. The results show that Ythdc2 inhibited the activity of reporter genes such as IFN1, NF-κB, IRF3, and ISRE commonly by affecting the activity of STING (Fig. 4E).
Fig. 4
Ythdc2 inhibits host antiviral innate immunity. A Relative protein and mRNA levels of Ythdc2 in MIC cells after transfected with NC or si-circYthdc2. B Relative protein levels of STING in MIC cells after transfected with NC or si-Ythdc2 and in MKC cells with pcDNA3.1 or Ythdc2. C and D qPCR assays were performed to determine the expression levels of IFN1, TNF-α, Mx1, ISG15, and Viperin in MIC cells transfected with NC or si-Ythdc2 (C) and transfected in MKC cells with pcDNA3.1 and Ythdc2 after SCRV infection (D). E Ythdc2 counteracts the negative effect of STING. Relative luciferase activities were detected in MKC after cotransfection with STING expression plasmid, pRL-TK Renilla luciferase plasmid, luciferase reporters, pcDNA3.1, Ythdc2. F Ythdc2 promotes SCRV replication. MIC cells transfected with NC or si-Ythdc2 and MKC cells were transfected with pcDNA3.1 or Ythdc2 for 24 h, respectively, then infected with SCRV at 24 h. The qPCR analysis was conducted for intracellular and supernatant SCRV RNA expression. (G) MIC cells seeded in 48-well plates overnight were treated with cultural supernatants at the dose indicated for 48 h. Then, cell monolayers were fixed with 4% paraformaldehyde and stained with 1% crystal violet. MIC cells were transfected with NC or si-Ythdc2. H Effect of Ythdc2 on cell viability after SCRV infection. MIC cells was transfected with NC or si-Ythdc2 for 24 h and then treated with SCRV for 24 h. Cell viability assay were measured. All data represented the mean ± SE from three independent triplicated experiments. *, p < 0.05; **, p < 0.01
Furthermore, we examined the effect of Ythdc2 on SCRV replication to explore the biological significance of Ythdc2 in SCRV-induced host cells. Through qPCR detection of SCRV RNA levels, we observed that silencing of Ythdc2 significantly inhibited SCRV replication 24 h after SCRV infection, resulting in a decrease of approximately 2/3 in SCRV levels (left panel of Fig. 4F). In addition, the Ythdc2 overexpression significantly promoted SCRV replication after 24 h of SCRV infection, resulting in an approximately twofold increase in SCRV levels (right panel of Fig. 4F). In addition, SCRV replication was also monitored by the appearance of CPE (cytopathic effect) in MIC cells. As shown in Fig. 4G, Knockdown of the Ythdc2 produced less CPE in the cells. These results demonstrated that Ythdc2 can enhance SCRV replication. When we investigated the effect of Ythdc2 on the cell viability of MIC cells, we found that the silence of Ythdc2 significantly increased cell viability after 24 h of SCRV infection (Fig. 4H). Collectively, Ythdc2 could inhibit STING-mediated antiviral immune response by promoting the degradation of STING protein, and all the results show that Ythdc2 has the same function of inhibiting host antiviral immune response as Ythdc2-170aa.
Ythdc2-170aa and Ythdc2 both promoted K48 and K11-linked ubiquitination of STINGTo determine which way was used to promote STING degradation by Ythdc2-170aa and Ythdc2. Therefore, we first explored the effects of circYthdc2 and Ythdc2 on the STING protein. As shown in Fig. 5A, the results show that overexpression of circYthdc2 or Ythdc2 could promote STING protein degradation, while silence of circYthdc2 or Ythdc2 could inhibit STING protein degradation. To further explore how sting protein is degraded, we tried to use protease inhibitors to block the degradation of circYthdc2 and Ythdc2 on STING. The results are shown in Fig. 5B, the protease inhibitor MG132 can effectively prevent circYthdc2 and Ythdc2 from degrading STING protein. In addition, we found that both circYthdc2 and Linear-FLAG-Ythdc2-170aa could effectively block the degradation of STING by MG132, and this result suggests that Ythdc2-170aa, the translation product of circYthdc2, may play a role in degrading STING. These results indicated that Ythdc2-170aa or Ythdc2 may promote STING degradation through the proteasome pathway. Immunostaining with Flag-specific antibodies shows a very high degree of colocalization both between Linear-FLAG-Ythdc2-170aa and GFP-STING or Flag-Ythdc2 and GFP-STING transfected into MKC cells (Fig. 5C).
Fig. 5
Ythdc2-170aa and Ythdc2 both promoted K11 and K48-linked ubiquitination of STING. A MKC cells were transfected with Flag-Ythdc2 and Flag-circYthdc2 plasmids, the cells were treated with 10 μM CHX for a different time before immunoblot analysis was performed; MIC cells were silence Ythdc2 or circYthdc2, and the cells were treated with 10 μM CHX for a different time before immunoblot analysis was performed B MKC cells were transfected with Linear-Flag-Ythdc2-170aa or Flag-circYthdc2 or Flag-Ythdc2 plasmids, after 42 h, the cells were treated with DMSO or 10 μM MG132 for 6 h before immunoblot analysis was performed. C Flag-Ythdc2 and Linear-Flag-Ythdc2-170aa were cotransfected with GFP-STING into MKC cells. Immunofluorescence staining using anti-Flag was performed to show the Ythdc2 and STING or Ythdc2-170aa and STING cellular localization. Original magnification is 630. D Immunoprecipitation and immunoblot analysis of Flag-Ythdc2 or Linear-Flag-Ythdc2-170aa with Myc-STING, in EPC cells. IP, immunoprecipitation. E Upper panel: Schematic diagram of Ythdc2-△HELICc plasmid construction. Lower panel: Immunoprecipitation and immunoblot analysis of Flag-Ythdc2, Flag-Ythdc2-△HELICc or Linear-Flag-Ythdc2-170aa with Myc-STING, in MKC cells. IP, immunoprecipitation. F Schematic diagram of STING, STING-△TM, STING-△N, STING-△C plasmid construction. G Immunoprecipitation and immunoblot analysis of Flag-Ythdc2, with Myc-STING, Myc-STING-△TM, Myc-STING-△N, Myc-STING-△C in MKC cells. IP, immunoprecipitation. H Immunoprecipitation and immunoblot analysis of Linear-Flag-Ythdc2-170aa with Myc-STING, Myc-STING-△TM, Myc-STING-△N, Myc-STING-△C in MKC cells. IP, immunoprecipitation. I Coimmunoprecipitation analysis of STING ubiquitination in EPC cells transfected with Myc-STING or HA-ubiquitin-WT in the presence of control vector, Flag-Ythdc2 or Linear-Flag-Ythdc2-170aa expression plasmid. IP, immunoprecipitation. J Coimmunoprecipitation analysis of STING ubiquitination in EPC cells transfected with Myc-STING or HA-ubiquitin-WT in the presence of control vector, Flag-Ythdc2, or Flag-Ythdc2-△HELICc, or Linear-Flag-Ythdc2-170aa expression plasmid. IP, immunoprecipitation. K Coimmunoprecipitation analysis of STING ubiquitination in MKC cotransfected with Myc-STING, Flag-Ythdc2 or Linear-Flag-Ythdc2-170aa expression plasmid and HA-ubiquitin-WT, HA-ubiquitin-K11 or HA-ubiquitin-K48 plasmids. All data represented the three independent triplicated experiments
To confirm that Ythdc2-170aa and Ythdc2 target STING, we investigated the interaction between Ythdc2-170aa and STING or Ythdc2 and STING. When cotransfected into MKC cells, Flag-tagged Ythdc2 (Flag-Ythdc2) coimmunoprecipitated with Myc-tagged STING (Myc-STING), and the Flag-tagged Ythdc2-170aa (Linear-FLAG-Ythdc2-170aa) also coimmunoprecipitated with Myc-STING (Fig. 5D). Moreover, compared with Ythdc2, Ythdc2-170aa has stronger binding ability with STING (Fig. 5D). Since Ythdc2 and Ythdc2-170aa share a common domain HELICc, we want to know whether the HELICc domain plays a role in binding to STING. Therefore, we constructed the HCLICc domain deletion mutant of the Ythdc2 (Flag-Ythdc2-△HELICc) plasmid. Immunocoprecipitation assay showed that the binding ability of STING to Ythdc2 decreased significantly after the HELICc domain’s deletion. Then, we cotransfected the Flag-Ythdc2 or Linear-FLAG-Ythdc2-170aa plasmids with Myc-STING plasmid and HA-ubiquitin plasmid (Fig. 5E).
To further explore which region of STING Ythdc2-170aa and Ythdc2 mainly bind to, we constructed the STING deletion mutant plasmid (Fig. 5F). Immunocoprecipitation assay showed that Ythdc2 mainly binds to the region between amino acids 1 and 157 of STING protein (Fig. 5G), and Ythdc2-170aa also mainly binds to the region between amino acids 42 and 157 of STING protein (Fig. 5H). Coimmunoprecipitation experiments showed that STING ubiquitination was markedly increased both in the presence of Ythdc2 and Ythdc2-170aa expression plasmid (Fig. 5I). Then, we tested whether Ythdc2 without HELICc domain could continue to promote the ubiquitination modification of STING. The results showed that Ythdc2-△HELICc could also promote the ubiquitination of STING, but the ubiquitination effect was greatly reduced compared with Ythdc2 (Fig. 5J). To further investigate Ythdc2 and Ythdc2-170aa-mediated STING ubiquitination, we used mutants in which all lysine residues except K48 or K11 were replaced with arginine (HA-ubiquitin-K48 and HA-ubiquitin-K11). Ythdc2 and Ythdc2-170aa-mediated ubiquitination of STING was significantly increased in the presence of wild-type HA-ubiquitin (HA-ubiquitin-WT) or HA-ubiquitin-K48 or HA-ubiquitin-K11 (Fig. 5K). These data indicate that Ythdc2 and Ythdc2-170aa both mediate K48 and K11-linked ubiquitination, and the HELICc domain is the key to the interaction between Ythdc2 and STING.
N6-Methyladenosine(m6A) modification mediates circYthdc2 translation polypeptidesCurrent research shows that there are two main ways to mediate circRNA translation, one is mediated by IRES and the other is mediated by m6A modification. Since Ythdc2, the host gene of circYthdc2, is an m6A-modified read protein, it is difficult not to associate whether there is m6A modification on circYthdc2. Then, by prediction, we found two potential m6A modification sites in the UTR region of circYthdc2. Therefore, we constructed the two potential m6A site mutant plasmids of circYthdc2 (Fig. 6A). Then, we transfected the wild-type and m6A site mutant FLAG-circYthdc2 plasmids into MKC cells, respectively. The results showed that the circYthdc2-170aa polypeptide expressed by the wild-type FLAG-circYthdc2 plasmid was significantly higher than that of the m6A site mutant FLAG-circYthdc2 plasmid. This result suggests that m6A modification may indeed exist on circYthdc2. We constructed some plasmids of m6A modification-related genes, including METTL3, METTL14, METTL16, FTO, ALKBH5, YTHDF1, YTHDF3, EIF3A, and EIF4G2. Subsequently, cotransfection experiments were conducted to verify whether these m6A modification-related genes could affect the translation efficiency of circYthdc2. As shown in Fig. 6B, the results indicated that METTL3, METTL14, YTHDF1, YTHDF3, and EIF4G2 could significantly increase the expression of circYthdc2-170aa polypeptide and significantly reduce the protein level of STING. It was also found that FTO could significantly reduce the expression of circYthdc2-170aa polypeptide and significantly increase the protein level of STING. To further verify the above experimental results, we conducted a concentration gradient experiment. The experimental results showed that METTL3, METTL14, YHTDF1, and YTHDF3 could significantly promote the expression of polypeptides in wild-type FLAG-circYthdc2 plasmids, but could not promote the expression of polypeptides in m6A site mutant plasmids (Fig. 6C).
Fig. 6
N6-methyladenosine modification mediates circYthdc2 translation polypeptides. A Upper panel: Schematic diagram of Flag-circYthdc2-m6A-mut plasmid construction. Lower panel: MKC cells were transfected with Flag-circYthdc2 or Flag-circYthdc2-m6A-mut plasmids, after 48 h, the immunoblot analysis was performed. B Myc-STING and Flag-circYthdc2 were cotransfected into MKC cells with m6A modification-related genes, respectively, and then the protein levels of Myc-STING and Flag-circYthdc2 were detected. C Left panel: Myc-STING and Flag-circYthdc2 were cotransfected into MKC cells with METTL3 or METTL14, respectively, and then the protein levels of Myc-STING and Flag-circYthdc2 were detected. Middle panel: Myc-STING and Flag-circYthdc2 were cotransfected into MKC cells with FTO or YTHDF1, respectively, and then the protein levels of Myc-STING and Flag-circYthdc2 were detected. Right panel: Myc-STING and Flag-circYthdc2 were cotransfected into MKC cells with YTHDF3 or Ythdc2, respectively, and then the protein levels of Myc-STING and Flag-circYthdc2 were detected. D Relative RNA levels of circYthdc2 in MKC cells after transfected with pcDNA3.1, METTL3, METTL14, YTHDF1, YTHDF3, FTO, and Ythdc2, respectively. E The m6A level alteration of circYthdc2 upon METTL3 or FTO overexpression was examined by MeRIP-qPCR. MKC cells were transfected with vector or METTL3 or FTO plasmid for 48 h. F The level of circYthdc2 upon YTHDF1 or YTHDF3 or Ythdc2 overexpression were examined by RIP-qPCR. MKC cells were transfected with vector or YTHDF1 or YTHDF3 or Ythdc2 plasmid for 48 h. G The protein level of YTHDF1 or YTHDF3 or Ythdc2 was examined by RNA pulldown. MKC cells were transfected MS2-GFP, MS2-circYthdc2 or MS2-circYthdc2-m6A-mut with vector or YTHDF1 or YTHDF3 or Ythdc2 plasmid for 48 h. H Relative luciferase activities were detected in MKC after cotransfection with STING expression plasmid, pRL-TK Renilla luciferase plasmid, luciferase reporters, circYthdc2, METTL3, METTL14, FTO, YTHDF1, YTHDF3. I Coimmunoprecipitation analysis of STING ubiquitination in MKC cotransfected with Myc-STING, Flag-circYthdc2 or m6A modification-related genes expression plasmid and HA-ubiquitin-WT, HA-ubiquitin-K11 or HA-ubiquitin-K48 plasmids. All data represented the mean ± SE from three independent triplicated experiments. *, p < 0.05; **, p < 0.01
The expression of circYthdc2-170aa polypeptides was closely related to the level of STING protein, when the expression of circYthdc2-170aa polypeptides increased, STING would correspondingly decrease, significant negative correlation; while FTO and Ythdc2 could significantly inhibit the expression of polypeptide in wild-type circYthdc2 plasmid. Notably, the expression of STING will be significantly decreased when transfected with Ythdc2 or circYthdc2 alone, but the expression of STING will be significantly increased when cotransfected with Ythdc2 and circYthdc2 (Fig. 6C). Therefore, we suspect that when the RNA level or polypeptide level of circYthdc2 rises, Ythdc2 will preferentially act on circYthdc2 instead of promoting the degradation of STING. Then, we tested the effects of METTL3, METTL14, FTO, YTHDF1, YTHDF3, and Ythdc2 on the RNA level of circYthdc2. The results showed that METTL3, METTL14, FTO, YTHDF1 and YTHDF3 had no significant effect on the RNA level of circYthdc2, but we found Ythdc2 could significantly reduce the RNA level of circYthdc2 (Fig. 6D). Furthermore, we explored its effect on the methylation level of circYthdc2 by performing a MeRIP-qPCR assay. Our results showed that overexpression of METTL3 dramatically increased the m6A level of circYthdc2 mRNA (Fig. 6E); in contrast, the m6A levels of circYthdc2 were significantly decreased after overexpression of FTO (Fig. 6E). To further verify whether YTHDF1, YTHDF3 and Ythdc2 can directly interact with circYthdc2, we conducted relevant RNA immunoprecipitation experiments. The experimental results showed that YTHDF1, YTHDF3, and Ythdc2 can significantly enrich circYthdc2, of which YTHDF1 has the highest enrichment efficiency, while Ythdc2 has a lower enrichment efficiency (Fig. 6F). At the same time, we also conducted RNA pulldown experiments. The results showed that wild-type circYthdc2 could enrich YTHDF1, YTHDF3 proteins, while m6A site mutant circYthdc2 could not enrich YTHDF1, and YTHDF3 proteins (Fig. 6G). Based on the above results, we believe that circYthdc2 can mediate the translation of polypeptides through m6A modification. METTL3 and METTL14 can act as “writer” proteins to promote the m6A modification of circYthdc2, FTO can act as “eraser” proteins to demethylate circYthdc2, and YTHDF1 and YTHDF3 can act as “reader” proteins to promote the translation of circYthdc2 into polypeptides, while Ythdc2 can act as a “reader” protein to promote the RNA degradation of circYthdc2.
We examined the effects of METTL3, METTL14, FTO, YTHDF1, and YTHDF3 on the NF-κB and IRF3 pathways. The results showed that METTL3, METTL14, YTHDF1, and YTHDF3 were roughly the NF-κB and IRF3 pathways that inhibited the activation of STING, while FTO was the NF-κB and IRF3 pathway that promoted the activation of STING (Fig. 6H). To further verify the above results, we cotransfected METTL3, METTL14, FTO, YTHDF1, YTHDF3, Ythdc2, and circythdc2 into MKC cells, respectively, and then detected the ubiquitination level of STING protein. The results were basically consistent with expectations. METTL3, METTL14, YTHDF1, and YTHDF3 could significantly promote the ubiquitination level of STING protein, while FTO and Ythdc2 could significantly inhibit the ubiquitination level of STING protein (Fig. 6I).
Ythdc2-170aa, the translation product of circYthdc2, which is highly conserved in structure and function in vertebratesThrough sequence alignment, we found that the sequence of exons 13 to 18 of Ythdc2 is highly conserved in amphibians, reptiles, birds, and mammals (including humans), all with a sequence length of 634 nt (Fig. 7A). Through further prediction, we speculate that exons 13 to 18 of Ythdc2 in these vertebrates can be reverse spliced to form circRNAs that can translate polypeptides. To confirm the objective existence of humans, Homo sapiens circYthdc2 (hsa-circYthdc2). First, hsa-circYthdc2 divergent primers were designed for RT-PCR amplification, and the amplified products were subjected to Sanger sequencing to confirm that hsa-circYthdc2 was spliced from the head to the tail (right panel of Fig. 7A). Through prediction, we found that the sequence of circYthdc2 is not only highly conserved in other vertebrates, but also may have a very conserved m6A modification site (Fig. 7B). Further studies found that circYthdc2 in different vertebrates may be able to translate polypeptides. We aligned the predicted amino acid sequences of circYthdc2 polypeptide, and the alignment results showed that the amino acid sequences of these polypeptides were highly conserved in vertebrates (Fig. 7C). To verify the correctness of the above prediction results, we constructed hsa-FLAG-circYthdc2, hsa-FLAG-circYthdc2-ATG-mut, and hsa-FLAG-circYthdc2-m6A-mut plasmids. Subsequently, the constructed plasmids were transfected into 293 cells, respectively, and the result showed that hsa-FLAG-circYthdc2 and hsa-FLAG-circYthdc2-m6A-mut could indeed be translated into polypeptide with about 24KD, while hsa-FLAG-circYthdc2-ATG-mut could not, and the expression level of the polypeptide translated by hsa-FLAG-circYthdc2 was much higher than that of hsa-FLAG-circYthdc2-m6A-mut. In addition, we also found that both hsa-FLAG-circYthdc2 and hsa-FLAG-circYthdc2-m6A-mut can promote the reduction of the level of human STING protein, while hsa-FLAG-circYthdc2-ATG-mut has no such effect (Fig. 7D). To further confirm that it can be driven by m6A modification, cotransfection experiments were conducted. As shown in Fig. 7E, the results indicated that METTL3, METTL14, and YTHDF1 could significantly promote the translation of polypeptide by hsa-FLAG-circYthdc2 plasmid and significantly reduce the protein level of STING, but not hsa-FLAG-circYthdc2-m6A-mut plasmid. It was also found that FTO could significantly reduce the translation of polypeptide by hsa-FLAG-circYthdc2 plasmid and significantly increase the protein level of STING, but not hsa-FLAG-circYthdc2-m6A-mut plasmid. In addition, we also found that silencing METTL3, METTL14, and YTHDF1 significantly increased the expression level of STING protein, while silencing FTO significantly reduced the expression level of STING protein (Fig. 7F). This result undoubtedly further confirms that circYthdc2 exists in humans and can be translated into polypeptide, and the inhibitory effect of its polypeptide on STING protein is similar to our confirmed results in teleost fish. Based on the above results, we believe that circYthdc2 is a highly conserved circRNA in vertebrates, and its translated polypeptides are also highly conserved. In addition, Ythdc2-170aa polypeptide has similar conserved functions both in fish and humans. The function of such a highly conserved circRNA must be of great significance.
Fig. 7
Ythdc2-170aa is highly conserved in structure and function in vertebrates. A CircYthdc2 exists in Miichthys miiuy, Nibea albiflora, Sciaenops ocellatus, Larimichthys ocellatus, Xenopus tropicalis (GenBank accession no. XM_031893156.1), Bufo gargarizans (GenBank accession no. XM_044275926.1), Podarcis muralis (GenBank accession no. XM_028748342.1), Numida meleagris (GenBank accession no. XM_021380759.1), Mus musculus (GenBank accession no. NM_001163013.1), Homo sapiens (GenBank accession no. NM_022828.5), and is composed of exon 13 to exon 18, with a length of 634nt. We confirmed the head-to-tail splicing of hsa-circYthdc2 in the hsa-circYthdc2 RT-PCR product by Sanger sequencing. B Sequence alignment of circYthdc2 from teleost fish to mammals. C Amino acid sequence alignment of circYthdc2 translated polypeptides from teleost fish to mammals. D HEK293 cells were transfected with vector or hsa-Flag-circYthdc2, hsa-Flag-circYthdc2-ATG-mut, hsa-Flag-circYthdc2-m6A-mut plasmids, after 48 h, the immunoblot analysis was performed. E HEK293 cells were transfected STING and hsa-Flag-circYthdc2 and hsa-Flag-circYthdc2-m6A-mut with METTL3 or METTL14 or YTHDF1 or FTO plasmids, after 48 h, the immunoblot analysis was performed. F HEK293 cells were transfected STING and hsa-Flag-circYthdc2 with si-has-METTL3 or si-has-METTL14 or si-has-YTHDF1 or si-has-FTO, after 48 h, the immunoblot analysis was performed. All data represented the three independent triplicated experiments
留言 (0)