
記住我
Correction to: J Mol Med (2016) 94:83–93 https://doi.org/10.1007/s00109-015-1327-6
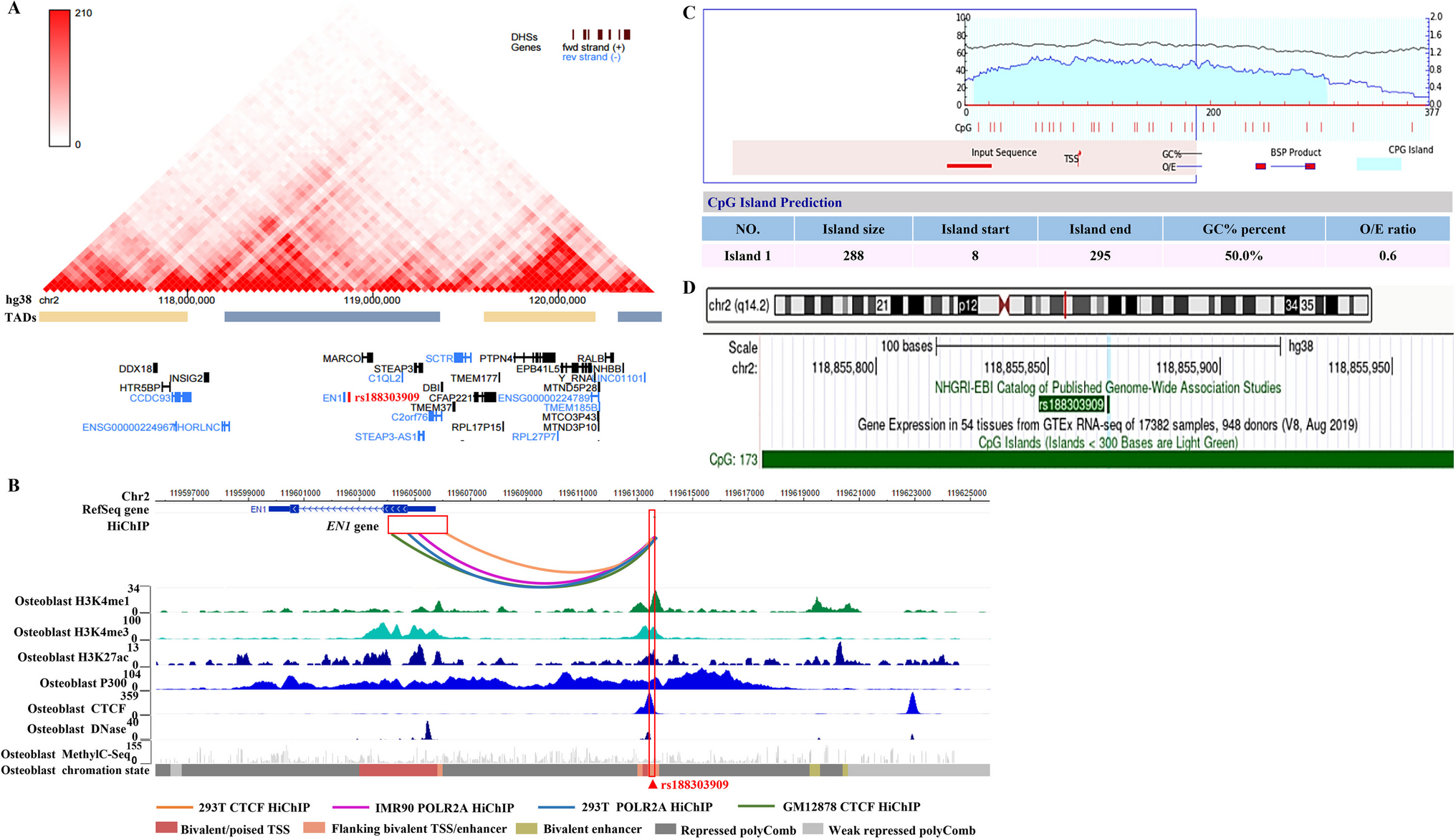
An unintended mistake occurred upon assembly of the images displayed in Fig. 5b, which show the separation of the target cells (CD19+ BV173 leukemia cells), effector cells (primary human NK cells: either unmodified or transduced with a chimeric antigen receptor (CAR) designed to target CD19) and clusters of effector and target cells that formed during co-culture experiments. These exemplary flow cytometry data plots show loss of CD19+ BV173 target cells and increased cell cluster formation in the case of co-culture with anti-CD19-CAR-NK cells (please compare target and cluster populations shown in the three exemplary flow cytometry data plots displayed in Fig. 5b of the original article). The effector populations in the three flow cytometry plots shown appear similar, however, closer inspection revealed that the pattern of many of the dots representing the NK cells appear identical in the plots named “Unmodified NK + BV173” (left-most flow cytometry data plot) and “CD19-CAR-NK + BV173” (middle flow cytometry data plot).
We have reviewed the flow cytometry data plots and data generated for the original paper and it appears that an error occurred when creating the exemplary plots of these co-culture experiments displayed in Fig. 5b. It seems that data from one condition were combined with data from a second condition. This resulted in the almost identical effector cell populations being displayed (blue dots), but differences in the target cell populations (red dots). Accordingly, different representations of the same displayed plot were created and included in Fig. 5b in the left (“Unmodified NK + BV173”) and middle (“CD19-CAR-NK + BV173”) images.
To repeat the experiments, primary human NK cells isolated from a new donor were modified with freshly prepared alpharetroviral vector supernatants for genetic transfer of the anti-CD19-CAR as previously done in the original work. The CD19+ target cell line BV173 was then cocultured with unmodified NK cells or with anti-CD19-CAR-NK cells with or without a CD19-blocking antibody following the same procedures as described in the original work. Analyses of these repeated experiments showed enhanced killing of the BV173 target cells upon coculture with the CAR-NK cells as compared to unmodified NK cells and that this enhanced killing of CD19+ BV173 cells by the anti-CD19-CAR-NK cells was reduced by co-administration of an anti-CD19 monoclonal antibody (please see data from repeated experiments in Fig. 5b and c below).
Thus, the conclusions drawn from the experiments shown in the original Fig. 5 and those from the repeated experiments are the same. Importantly, none of the main messages of the article were impacted by this error. However, as the data from the original Supplemental Fig. S3A was dependent on experimental data contained in the original Fig. 5c, the depiction of which can no longer be fully reconstructed, Supplemental Fig. S3A has been deleted the data shown in the original Supplemental Fig. S3B is now shown as Supplemental Fig. S3A.
These changes also impact the text in four instances in the manuscript. Please see the corrected text in the following paragraphs.
Corrections to this passage:
Cytotoxicity analyses with CAR transduced primary human NK cells
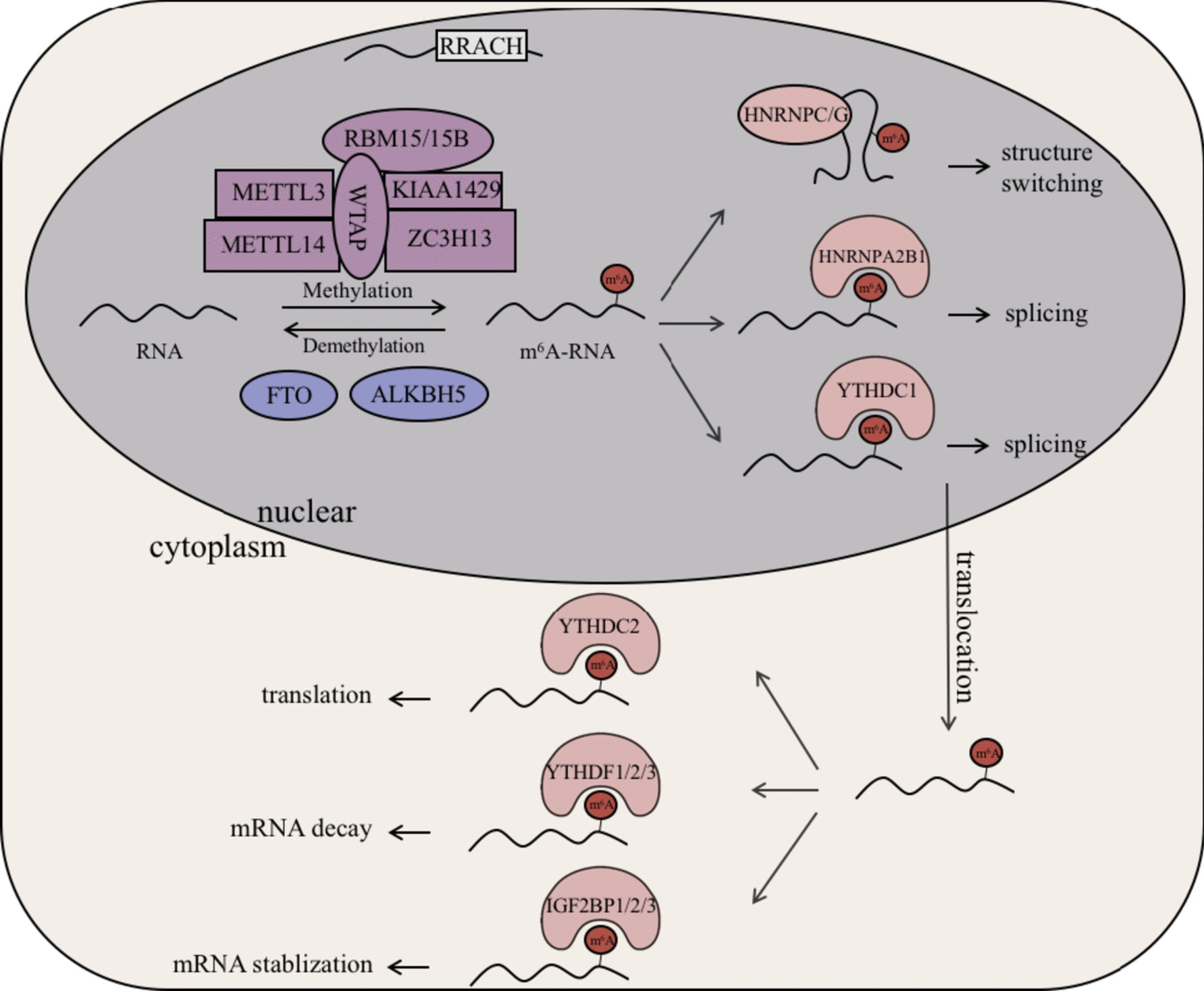
Tumor-specific cytotoxicity of NK cells can be enhanced by engineering NK cells to express CARs. We designed a codon-optimized anti-CD19 CAR, which consists of a signal sequence derived from granulocyte–macrophage colony-stimulating factor (GM-CSF) receptor α-chain, anti-CD19 scFv derived from a mouse hybridoma FMC63 [27], CD28, CD137 (4-1BB), and CD3ζ [28]. This codon-optimized anti-CD19 CAR was cloned into alpharetroviral SIN vectors, which, similarly to the previously used EGFP vectors, contained the MPSV promoter and the wPRE (Fig. 5a). Retronectin-mediated transduction of primary human NK cells was performed at MOIs 1 and 3 using RD114/TR-pseudotyped alpharetroviral anti-CD19 CAR particles. Transduced NK cells were coincubated with CD19-positive BV173 leukemia cells at different ratios and monitored for 48 h using a single platform, no-wash flow cytometric assay [30, 31]. Compared to unmodified primary NK cells, anti-CD19 CAR-modified NK cells exhibited improved tumor-specific cytotoxicity as shown by a decreased percentage of BV173 leukemia target cells (Fig. 5b, Supplementary Fig. S2). This assay allowed absolute quantification of viable leukemic cells and revealed an enhanced tumor-specific cytotoxicity upon comparison of transduced and untransduced NK cells (Fig. 5c, Supplementary Fig. S3). BV173 cells cultivated in the absence of NK cells expanded more than threefold over the course of these experiments (data not shown). Interestingly, high effector/target ratio led to a nearly complete lysis of CD19+ BV173 cells after 48 h, which was blocked using anti-CD19 antibodies (Fig. 5c).
Corrections to Fig. 5 legend:
Fig. 5 Cytotoxicity assay. a Schematic depiction of the alpharetroviral SIN vector used in this study. The long-terminal repeat (LTR) (ΔU3, R, and U5), packaging signal (ψ), myeloproliferative sarcoma virus (MPSV) promoter, signal peptide (SP), CD19 single-chain variable fragment (CD19 scFv), and woodchuck hepatitis virus post-transcriptional regulatory element (wPRE) are indicated. b Exemplary flow cytometry plots of different samples at 18 h after coincubation of primary NK cells with BV173 leukemia cells. Presence of leukemia target cells based on CD56-PC7 and CD9-FITC detection in mixtures of untransduced/ transduced NK cells (CD56-positive) with BV173 leukemia target cells (CD9-positive and CD19-positive) at a ratio of approximately 0.5:1 (effector/target). Addition of anti-CD19 antibodies (mAB) inhibited anti-leukemic activity of CD19 CAR-modified NK cells, as indicated by higher numbers of BV173 cells in cocultures containing anti-CD19 mAB. As BV173 leukemia target cells express CD9 and CD19, BV173 leukemia cell numbers were monitored with a CD9-FITC antibody to avoid possible interaction between the CD19scFv and the CD19 blocking antibody. c Coincubation of NK cells with leukemia target cells at a ratio of 5:1 (effector/target) (left panels) or 0.5:1 (effector/target) (right panels). The change in numbers of leukemia cells over time is expressed in percent relative to the leukemia cell numbers initially present (t = 0.1 h) in cocultures containing untransduced NK cells. Multiplicities of infection (MOIs) were determined on human fibroblast cells (HT1080). Shown are mean values + SD, from one experiment performed in duplicate with NK cells from one donor. Additional experiments at higher MOI and using different virus preparations to transduce NK cells from another NK cell donor are depicted in Supplementary Fig. S3A
Corrections to page 91 of original article:
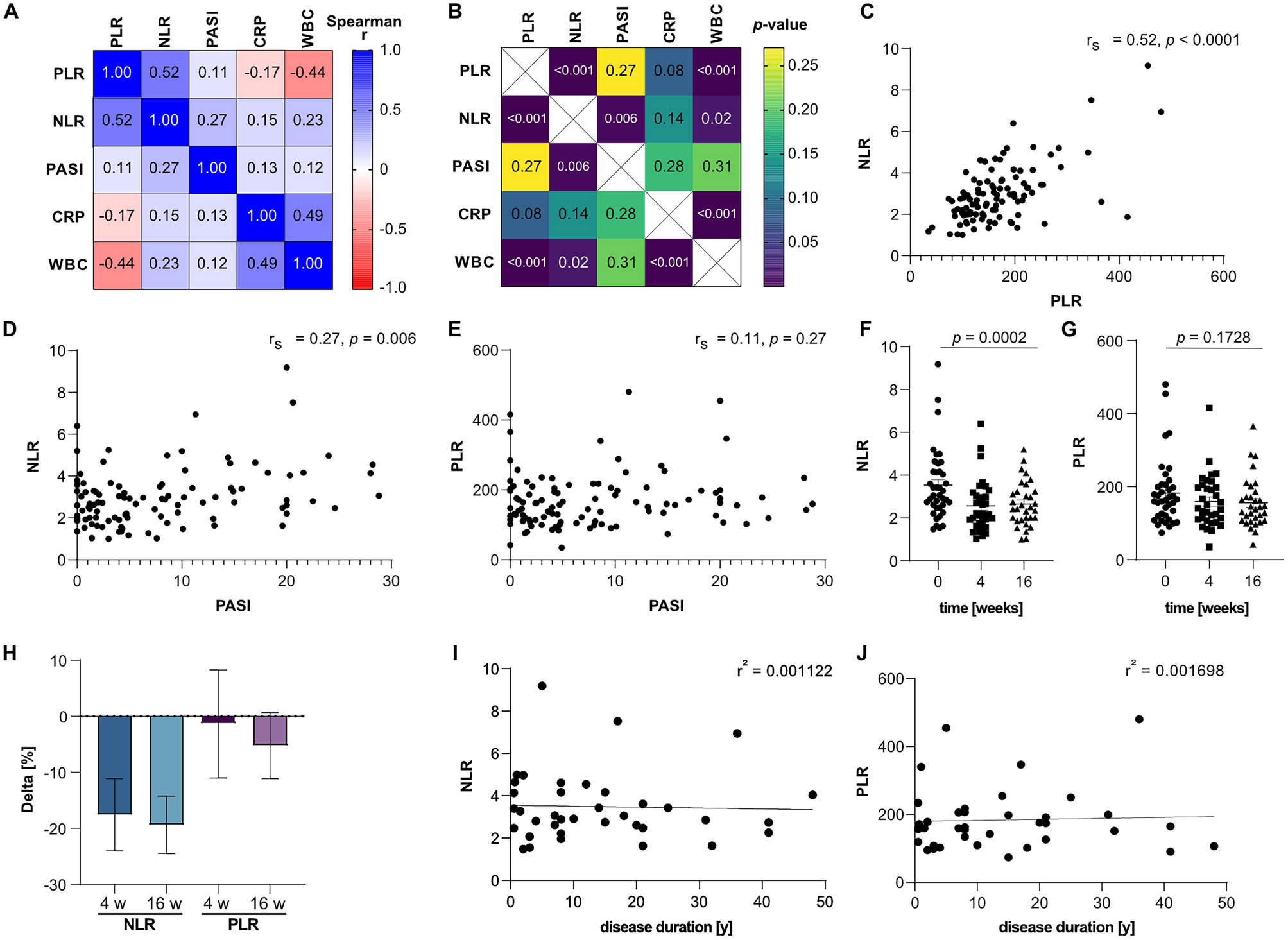
Combining the alpharetroviral vector platform for efficient NK cell transduction with improved NK cell expansion protocols might represent important progress to clinical development of NK cell-based therapies. Modified NK cells could be administered to improve current therapeutic strategies, such as in combination with chemotherapy or with other targeted therapies. Use of third-party NK cells might provide clinical benefit when combined with haploidentical allogeneic transplantation in heavily treated patients. The relatively short half-life of NK cells may allow improved temporal delivery of desired effects, such as targeted cytotoxicity by modification with tumor antigen specific CARs. Previous clinical NK cell studies showed mature donor NK cells persisted in vivo for 1– 3 weeks following haploidentical stem cell transplantation or third-party donor NK cell infusion [8, 9]. It may be possible to exploit distinct characteristics of NK cells from different donors as these may vary in capacities such as cytotoxicity (compare untransduced cells in Fig. 5c and Supplementary Fig. S3A). An interesting therapeutic modality would be to combine NK cell and T cell infusion to deliver a more rapid short-term response concomitantly with a long-lasting response.
Correction to Original Legend to Supplementary Fig. S3:
Supplementary Fig. S3 Coincubation of CD19 CAR-modified NK cells with leukemia target cells (CD19+ BV173 cells). The change in numbers of leukemia cells over time is expressed in percent relative to the leukemia cell numbers initially present (t = 0.1 h) in cocultures containing untransduced NK cells. (A) Depicted are the cytotoxicities of NK cells isolated from another donor, whose NK cells exhibit slightly different cytotoxicity characteristics than NK cells from the donor whose cells were used in Fig. 5c. These NK cells were also modified with the same CD19 CAR construct (depicted in Fig. 5a) and used in Fig. 5c. In these experiments, NK cells were coincubated with BV173 cells at a ratio of approximately 5:1 (effector to target ratios) (left panels) or 0.5:1 (effector:target) (right panels) in 5 mL polystyrene round bottom snap cap tubes (BD, Heidelberg, Germany) in CellGro medium (CellGenix, Freiburg im Breisgau, Germany) supplemented with 5% human AB serum, 1000 I.E./mL Proleukin S (Novartis Pharma, Nuremberg, Germany) under constant shaking at 37 °C, 5% CO2. For analysis, CD19 PE (clone HIB19, eBioscience, Frankfurt am Main, Germany), CD9 FITC (clone ALB6, Beckman Coulter, Krefeld, Germany), CD56 PerCPVio700 (clone REA196, Miltenyi, Bergisch-Gladbach, Germany) antibodies were added, followed by incubation at 4 °C in the dark for 30 min. CountBright Absolute Counting Beads (Molecular Probes, Life Technologies, Darmstadt, Germany) and DAPI were then added followed by immediate flow cytometric data acquisition on the BD LSRII flow cytometer (BD Biosciences, Heidelberg, Germany). Data analyses were performed with FACSDiva software (BD Biosciences, Heidelberg, Germany). Shown are results for one measurement for untransduced NK cells and NK cells transduced with an MOI of 1, and for duplicate measurements (+SD) with cells transduced with an MOI of 3 from one experiment performed with NK cells from one donor.
All authors approved the correction to this article and wish to apologize for any inconvenience caused by this mistake.


Corrected Supplementary Fig. S3:

留言 (0)