
Remember me
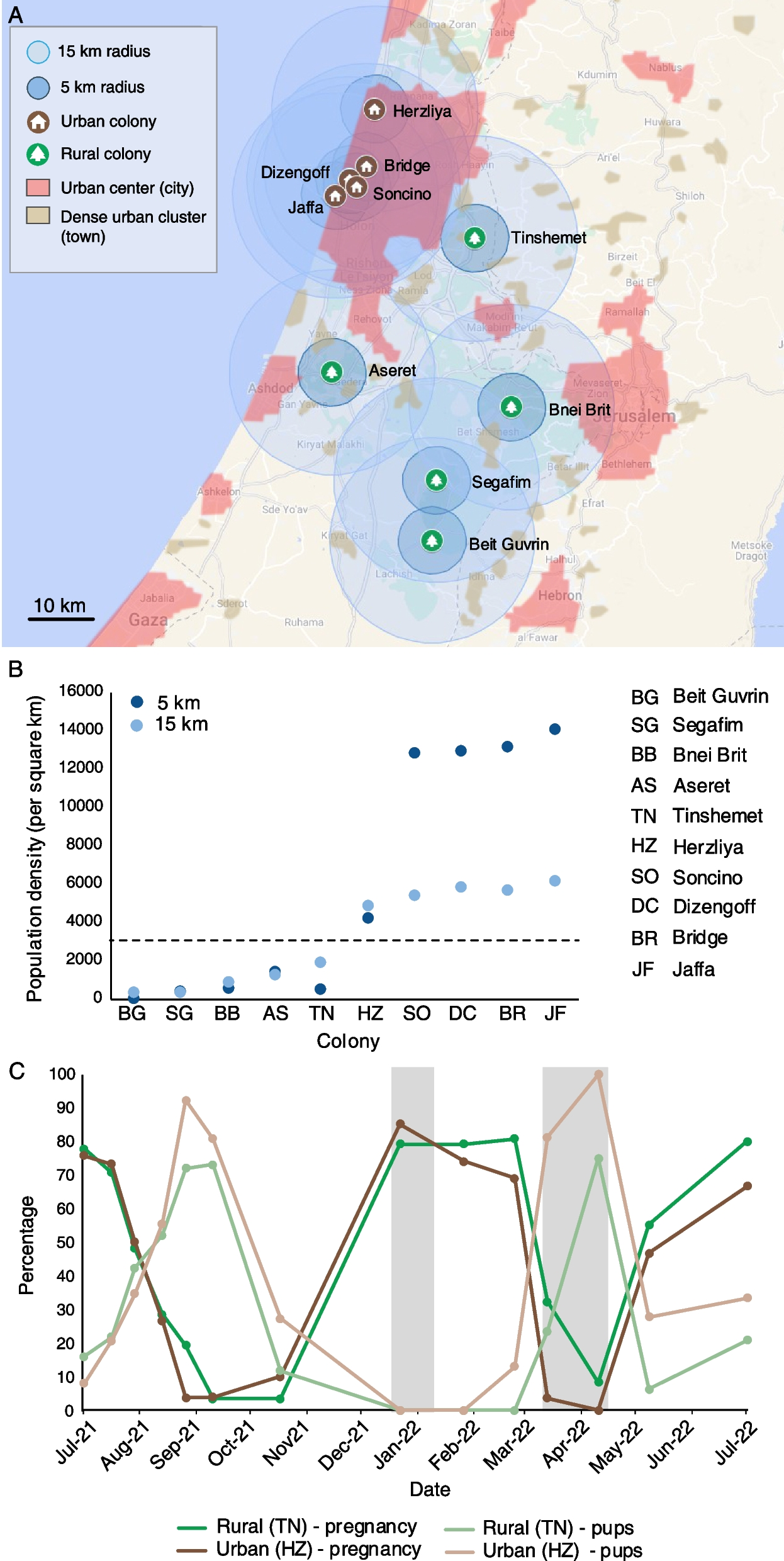
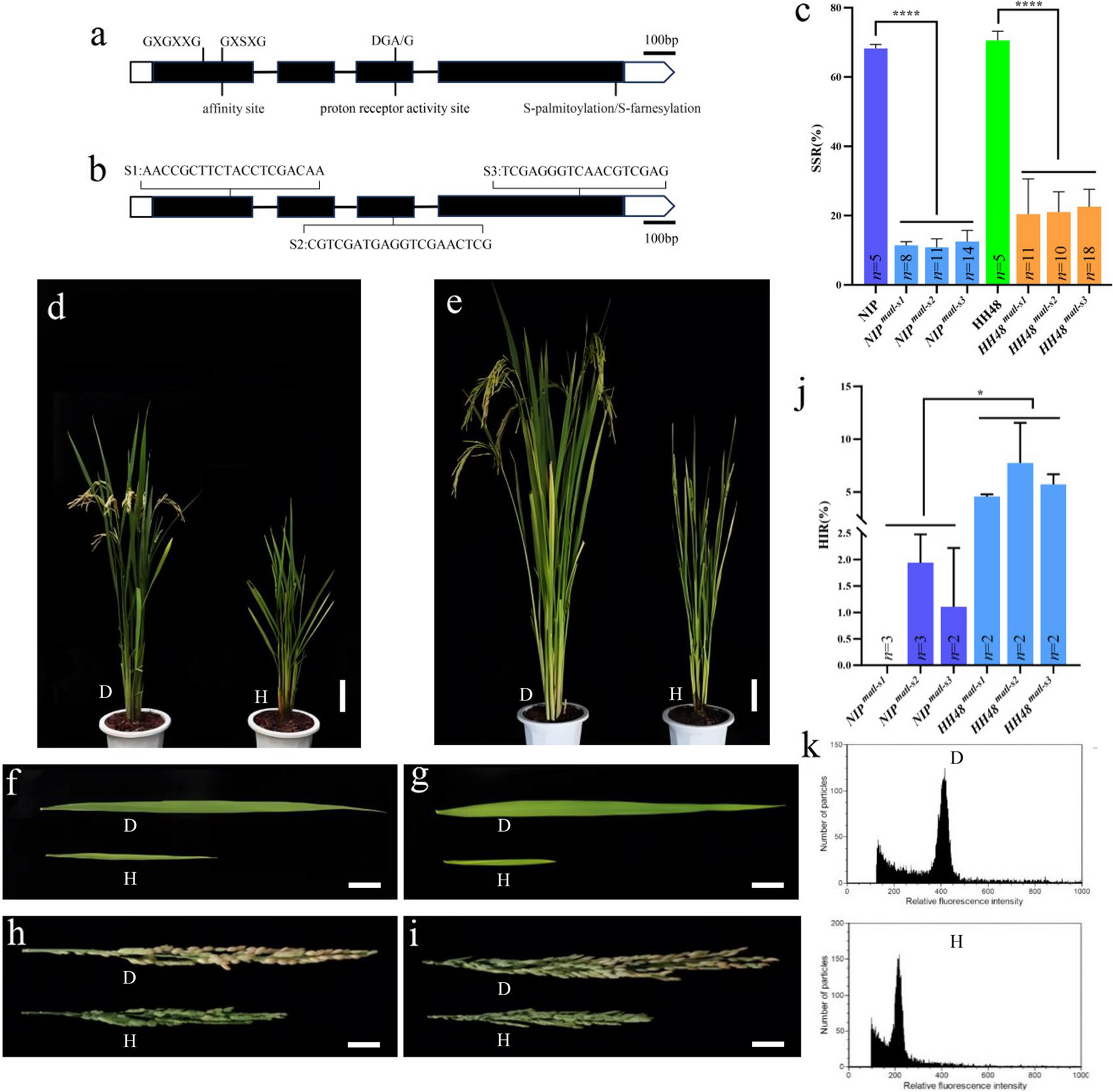





Loss of Fgf9 in the developing testis leads to somatic sex reversal and consequent germline sex reversal. To avoid this significant confounding factor, we allowed sex determination to occur before inhibiting FGFR or MEK1/2 and assessed how this affected male germline differentiation in an intact gonad. This allowed us to temporally separate somatic sex determination from male-specific germline differentiation. To achieve this, E12.5 XY gonad-mesonephros samples were cultured with vehicle control (DMSO) or a range of small molecule inhibitors of FGFR1-3, MEK1/2, p38MAPK, and PI3K to screen for germline developmental effects mediated by these signalling pathways. Inhibitors initially used included pan-FGFR (FGFR1-3) inhibitor BGJ398/Infigratinib (FGFRi; IC50 ~ 1.0 nM), MEK1/2 inhibitor PD0325901/Mirdametinib (MEKi; IC50 0.33 nM), p38MAPK inhibitor PH-797804 (p38i; IC50 26 nM) or PI3K inhibitor GSK1059615 (PI3Ki, IC50 5 nM). Importantly, all of these inhibitors have IC50 values in the low nM range and have been extensively validated in clinical trials demonstrating their high specificity, potency and cell tolerance (Table 1).
Table 1 Summary of treatments and doses used in gonad culturesTo account for the difference between cell free IC50 values and drug bioavailability in gonad culture, we initially used a starting concentration of 500 nM for each drug. This is consistent with our observation that drugs with similar IC50s maximally inhibit their targets in gonad cultures in the 100–1000 nM range. The vehicle control, DMSO, was used at a dilution of ≥ 1/5000 in all experiments, a concentration that does not affect gonad or germline development [20, 38]. Bright-field and fluorescence examination of E12.5 testis-mesonephros samples cultured with control (DMSO) or drug for 72 h provided an initial readout of the impact of each drug based on germ cell organisation within testis cords, marked by germ cell-specific expression of Oct4-eGFP (Fig. 1A). DMSO controls developed well-defined cords containing germ cells, but FGFRi and MEKi resulted in poor testis cord structure and some GFP-positive germ cells were located outside the testis cords. p38i- and PI3Ki-treated gonads were morphologically similar to DMSO controls, with GFP-positive germ cells contained within well-defined testis cords (Fig. 1A).
Fig. 1
FGF and MEK1/2 inhibition disrupts foetal testis development but only MEK1/2 inhibition disrupts germ cell mitotic arrest. A Bright-field and GFP images of E12.5 XY gonad-mesonephros tissue cultured for 72 h with DMSO, or 500 nM of FGFRi, MEKi, p38i or PI3Ki. Scale bar: 100 μm. Dotted lines highlight the gonad. B–E Flow cytometric analysis of Sertoli (B,D) or germ (C,E) cell proliferation based on EdU incorporation in XY E12.5 gonad-mesonephros tissue cultured for 72 h with DMSO, 500 nM of FGFRi, MEKi, PI3Ki or p38i (B,C) or 125, 250, 500 or 1000 nM of FGFRi or MEKi (D,E). F,G Flow cytometric analysis of Sertoli (F) or germ (G) cell proliferation based on EdU incorporation in XY E13.5 gonad-mesonephros tissue cultured for 48 h with DMSO or 500 nM of FGFRi or MEKi. H Immunofluorescent images of E12.5 gonad-mesonephros tissue cultured for 24 h with DMSO, 500 nM of FGFRi or MEKi demonstrating MEK1/2 signalling activity. Top panel: DAPI (blue), MVH (green), NR2F2 (red), pERK1/2 (cyan). Bottom panel: pERK1/2 (grey). Scalebar represents 50 μm. Replicates: A–C n = 6–9, D,E n = 3–16, F,G n = 3, H n = 3–4. Statistics: B, D, E Ordinary one-way ANOVA with Tukey’s multiple comparison, C Brown-Forsythe and Welch ANOVA with Dunnett’s T3 multiple comparisons, F,G Unpaired two-tailed t-test. Error bars: Mean ± SEM. Significance between control and treatment: *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
To further determine the effects of each treatment on testis development, we added 5-ethynyl-2’-deoxyuridine (EdU) during the final 2 h of culture and used flow cytometry to assess proliferation based on EdU incorporation during S-phase and DNA content using propidium iodide (PI) staining in E12.5 gonad-mesonephric samples cultured for 72 h with each inhibitor [20, 38, 39] (Fig. 1B,C). As expected, Sertoli cells were highly proliferative in DMSO controls (Fig. 1B). However, FGFRi or MEKi reduced Sertoli cell proliferation compared to XY DMSO controls (P < 0.05 or P < 0.001, respectively, Fig. 1B). As expected, germ cell proliferation was very low in DMSO controls (Fig. 1C), confirming that the germ cells had entered mitotic arrest, a key milestone in male germline differentiation [2]. In contrast, germ cell proliferation was substantially higher in MEKi-treated gonads (P < 0.05), demonstrating that germ cell mitotic arrest was disrupted (Fig. 1C). However, the percentage of germ cells incorporating EdU in FGFRi-treated testes was similar to DMSO controls, demonstrating that the germ cells had entered mitotic arrest (Fig. 1C). As FGFRi was expected to disrupt germ cell mitotic arrest, this outcome was confirmed using another potent FGFR inhibitor (AZD4547; Table 1), which also resulted in reduced Sertoli cell proliferation but did not disrupt germ cell mitotic arrest (Additional file 1: Fig. S1A,B). Neither p38i (PH-797804) nor PI3Ki (GSK1059615) affected Sertoli cell proliferation or germ cell mitotic arrest (Fig. 1B,C), an outcome confirmed using independent, p38 and PI3K inhibitors, ralimetanib dimesylate (LY2228820) and PF-04691502 (Table 1; Additional file 1: Fig. S1C,D).
Given both FGFRi and MEKi reduced Sertoli cell proliferation and disrupted testis cord development, we analysed AMH, SOX9 and FOXL2 using immunofluorescence (IF) to ensure that FGFR or MEK inhibition did not result in somatic sex reversal. Notably, robust SOX9 and AMH staining was detected by IF in male DMSO, FGFRi and MEKi samples but not in female gonads (Additional file 2: Fig. S2A), indicating that treated XY gonads maintained a male phenotype. Consistent with this, assessment of SOX9 intensity using flow cytometry demonstrated that SOX9 expression was not reduced in FGFRi- or MEKi-treated samples compared to DMSO controls (Additional file 2: Fig. S2B). Furthermore, IF staining for the female marker, FOXL2, revealed strong expression in female gonads, but minimal staining in male DMSO, FGFRi or MEKi samples, although occasional FOXL2-positive cells were detected in MEKi-treated samples (Additional file 2: Fig. S2C). Importantly, these data demonstrate that inhibition of FGFR or MEK1/2 at E12.5 did not result in somatic sex reversal. This allowed us to further assess the effects of FGF or MEK1/2 inhibition in the environment of a developing testis on male germline differentiation and in the absence of somatic sex reversal.
FGF and MEK1/2 signalling are both required for Sertoli cell proliferation, but only MEK1/2 is required for germ cell mitotic arrestTo titrate the dose response to MEK1/2 and FGFR inhibition, E12.5 XY gonad-mesonephros samples were cultured with MEKi or FGFRi at 0 (DMSO diluted ≥ 1/5000), 125, 250, 500 and 1000 nM and assessed using flow cytometry (Fig. 1D,E and Additional file 3: Fig. S3). Compared to DMSO control, proliferation of SOX9 expressing Sertoli cells was reduced by all doses of FGFRi ≥ 125 nM (P < 0.0001; Fig. 1Di) and ≥ 250 nM MEKi (P < 0.0001, Fig. 1Dii). Of interest, although the maximal impact of FGFRi on Sertoli cell proliferation occurred at 125 nM, it did not further reduce Sertoli cell proliferation even at 1000 nM and this effect was noticeably less than that of MEKi at doses of 500 and 1000 nM (Fig. 1Di vs Dii). Based on these data, doses of 500 nM of FGFRi and 500 nM of MEKi were used for further experiments.
Consistent with our initial observations (Fig. 1C), although Sertoli cell proliferation was reduced, germ cell mitotic arrest remained unaffected by FGFRi even with a dose of 1000 nM (Fig. 1Ei). To confirm this outcome, we tested whether a very high dose of 2500 nM FGFRi, which is ~ 2500 × the IC50 value (Table 1) and 20 × the 125 nM minimal dose affecting Sertoli cell proliferation (Fig. 1Di), might affect germ cell mitotic arrest. However, this again resulted in a similar reduction in Sertoli cell proliferation, but no effect on germ cell mitotic arrest (Additional file 1: Fig. S1E-F). We next compared outcomes for FGFRi and AZD4547 with and third inhibitor, SU5402, which was previously used at 5000 nM to inhibit FGFR [9]. Confirming the outcomes obtained using FGFRi and AZD4547, 5000 nM SU5402 reduced Sertoli cell proliferation to a similar extent as FGFRi and AZD4547, but it did not affect germ cell mitotic arrest (Table 1; Additional file 1: Fig. S1E-F). In contrast, MEKi potently disrupted germ cell mitotic arrest even when used at 125 nM, with increasingly high proportions of EdU-positive proliferative germ cells as MEKi concentration increased (P < 0.0001, Fig. 1Eii).
In repeated experiments, MEK1/2 inhibition profoundly disrupted germ cell mitotic arrest in E12.5 XY gonads but three different FGFR inhibitors did not, even though FGFs typically elicit a response through MEK1/2-pERK1/2 within 10–15 min [25, 26]. As FGF9 expression peaks at E11.5 [11], a potential explanation for the inability of FGFRi to disrupt germ cell mitotic arrest could be that FGF9 was inhibited too late in E12.5 cultures. However, as Fgf9 or Fgfr2 genetic deletions cause somatic sex reversal [6, 7, 12,13,14], inhibition of FGFR at E11.5 is expected to cause somatic sex reversal and consequent germ cell sex reversal that would substantially confound data interpretation. To avoid the confounding effect of male to female sex reversal and provide a clearer outcome for the study, we did not include studies of FGFR inhibition at E11.5.
To determine if MEK1/2 inhibition affected Sertoli cell proliferation and germ cell mitotic arrest after E13.5, E13.5 XY gonad-mesonephros samples were cultured for 48 h with DMSO or 500 nM MEKi. MEKi treatment from E13.5 significantly reduced Sertoli (P < 0.01, Fig. 1F) and increased germ cell proliferation (P < 0.01, Fig. 1G). However, the effect of MEKi on germ cell mitotic arrest was diminished compared to E12.5 (E13.5: EdU 6% vs E12.5: 15%, P < 0.0001 E12.5 MEKi vs E13.5 MEKi; Fig. 1Eii,G), indicating that the ability of MEKi to disrupt mitotic arrest decreased between E12.5 and E13.5.
We previously demonstrated that FGF9 induces proliferation of XX somatic cells at rates similar to XY gonads [20]. To confirm that FGFRi and MEKi effectively blocked FGF9 activity, XX E12.5 gonads were cultured for 48 h in media containing DMSO, 50 ng/mL FGF9, 500 nM FGFRi, 500 nM MEKi, FGF9 + FGFRi or FGF9 + MEKi and assessed using flow cytometry. As expected, FGF9 substantially increased the proliferation of XX gonadal somatic cells compared to XX controls (P < 0.05, P < 0.0001 ; Additional file 1: Fig. S1G). Critically, both FGFRi and MEKi completely neutralised FGF9, with somatic cell proliferation decreased to XX control levels in FGF9 + FGFRi or FGF9 + MEKi-treated XX gonads (Additional file 1: Fig. S1Gi,Gii). In contrast, neither p38i nor PI3Ki counteracted FGF9-induced somatic cell proliferation in XX gonads, indicating that neither p38MAPK nor PI3K regulate primary pathways through which FGF drives somatic cell proliferation in developing gonads (Additional file 1: Fig. S1Giii).
Inhibition of MEK1/2 completely abolished ERK1/2 phosphorylation in the developing testis, but FGFR inhibition did notFGF activation of MEK1/2 rapidly results in phosphorylation of ERK1/2, and MEK1/2 inhibition blocks this activity [25, 26]. To determine if inhibition of FGF or MEK1/2 signalling prevented phosphorylation of ERK1/2, E12.5 XY gonad-mesonephros samples were cultured with 500 nM FGFRi or MEKi for 24 h and pERK1/2 was assessed using IF (Fig. 1H and Additional file 1: Fig. S1H). Surprisingly, pERK1/2 was not detected in MVH (mouse vasa homolog) expressing germ cells in the developing testis. However, consistent with MEK1/2 activity in Sertoli cells, pERK1/2 was detected at low levels in MVH negative somatic cells within testis cords in DMSO controls. While there are no other somatic cells within testis cords, Sertoli cell localisation of pERK1/2 could not be definitively determined using SOX9 IF as the SOX9 and pERK1/2 antibodies were both raised in rabbit. Robust pERK1/2 was also detected in somatic cells outside of the testis cords that appeared to be endothelial cells; however, this was not confirmed. pERK1/2 was not detected in MEKi-treated samples, although it was detected in Sertoli and somatic cells outside of the testis cords in FGFRi-treated gonads (Fig. 1H and Additional file 1: Fig. S1H) demonstrating that MEKi abolished ERK1/2 phosphorylation, but FGFRi did not. Consistent with this, MEKi reduced Sertoli cell proliferation to a greater extent than FGFRi (P < 0.0001 MEKi vs FGFRi, Additional file 1: Fig. S1I). However, as 500 nM of FGFRi completely blocked FGF9-induced proliferation in XX somatic cells (Additional file 1: Fig. S1Gi), the most likely explanation for the inability of FGFRi to completely eliminate pERK1/2 in Sertoli cells is that MEK1/2 may be activated independently of FGFR, perhaps by PGD2 [1, 8, 40] or other ligands.
FGF and MEK1/2 signalling is required for normal testis cord formationAs gonad whole-mount images indicated testis cords were disrupted by FGFRi or MEKi (Fig. 1A), we used IF to investigate MVH-positive germ cells relative to SOX9 expressing Sertoli cells, SMA (smooth muscle actin) expressing peritubular myoid cells or laminin, which delineate testis cords. In DMSO controls, the majority of Sertoli cells were organised in a single layer at the testis cord basement membrane, with germ cells very rarely found outside the cords (Fig. 2). In FGFRi- and MEKi-treated samples, some Sertoli cells localised to the testis cord basement membrane, but gaps were evident between the Sertoli cells, and many Sertoli cells remained dispersed throughout the interior of the testis cords (Fig. 2A). Furthermore, germ cells were occasionally present in the gaps between Sertoli cells at the testis cord basement membrane (Fig. 2A) and were mis-localised outside testis cords in FGFRi-treated cultures, although this was more common in MEKi-treated gonads (Fig. 2B).
Fig. 2
FGF and MEK1/2 signalling is required for normal testis cord formation. Immunofluorescent images of XY E12.5 gonad-mesonephros tissue cultured with DMSO, 500 nM FGFRi or 500 nM MEKi for 72 h showing Sertoli (A) and germ (A,B) cell localisation. DAPI (blue), MVH (green), SMA (red: A) or Laminin (red: B) and SOX9 (cyan: A). Scale bars: top panel 100 μm, bottom panel 50 μm. Replicates: n = 3–4. A White arrows identify gaps in the Sertoli cell layer at the testis cord basement membrane; white asterisks identify Sertoli cells dispersed within the inner area of the testis cords. B White arrows identify germ cells localised outside testis cords
MEK1/2 signalling is required for male germline differentiationAs MEKi prevented germ cell mitotic arrest, the expression of male germline differentiation markers was assessed using IF and flow cytometry in E12.5 XY gonad-mesonephros samples cultured for 72 h with DMSO, 125 or 500 nM FGFRi or MEKi. DPPA4 is expressed in XX and XY germ cells at E12.5, but is upregulated in XY germ cells and repressed in XX germ cells as they differentiate [20]. As expected, DPPA4 was not detected in germ cells of XX gonads but was detected in germ cells of XY E12.5 gonads cultured for 72 h with DMSO, and fluorescence appeared more intense in XY E12.5 + 72 h than in E12.5 XY germ cells (Fig. 3A and Additional file 4: Fig. S4A). While DPPA4 germ cell levels were similar in XY E12.5 + 72 h DMSO and FGFRi cultures, DPPA4 intensity appeared lower in MEKi-treated samples and comparable to E12.5 XY germ cells (Fig. 3A and Additional file 4: Fig. S4A). Confirming this, flow cytometry revealed that the relative DPPA4 germ cell intensity was 2 × higher in XY E12.5 + 72 h DMSO and FGFRi cultures than in E12.5 XY germ cells (P < 0.0001), but DPPA4 was expressed at comparable levels in MEKi-treated samples compared to E12.5 XY germ cells (Fig. 3B).
Fig. 3
MEK1/2 signalling is required for male germline differentiation. Analysis of E12.5 XY gonad-mesonephros or E12.5 XY or XX gonad-mesonephros cultured for 72 h with DMSO, 125 or 500 nM FGFRi or MEKi. A Immunofluorescent images demonstrating DPPA4 localisation. Top panel: DAPI (blue), MVH (green), DPPA4 (red), SMA (cyan). Bottom panel: DPPA4 (grey). B DPPA4 staining intensity in germ cells determined by flow cytometry. C Immunofluorescent images demonstrating DNMT3L localisation. Top panel: DAPI (blue), MVH (green), NR2F2 (red), DNMT3L (cyan). Bottom panel: DNMT3L (grey). D,E Percentage DNMT3L-positive germ cells (D) and DNMT3L staining intensity (E) determined by flow cytometry. F Immunofluorescent images demonstrating PIWIL2 localisation. Top panel: DAPI (blue), MVH (green), SMA (red), PIWIL2 (cyan). Bottom panel: PIWIL2 (grey). Scale bars: 50 μm. Replicates: A, C, F n = 3–4, B n = 4–14, D,E n = 4–9. Statistics: B,E Brown-Forsythe and Welch ANOVA with Dunnett’s T3 multiple comparisons, D Ordinary one-way ANOVA with Tukey’s multiple comparison. In B,E, Intensity is relative to E12.5 + 72 h XY DMSO control sample set at 1.0. Error bars: mean ± SEM. Significance between control and treatment: **P < 0.01, ***P < 0.001, ****P < 0.0001
As germ cell proliferation and DPPA4 levels indicated that male germ cells failed to properly differentiate in MEKi-treated XY gonads, we examined two additional male germline markers, DNMT3L and PIWIL2. IF and flow cytometry revealed that the majority of germ cells were DNMT3L positive in DMSO controls and FGFRi-treated gonads (Fig. 3C–E and Additional file 4: Fig. S4B). In contrast, very few germ cells were DNMT3L positive in MEKi-treated gonads (P < 0.01, P > 0.0001), and the DNMT3L staining intensity was significantly lower than in the DMSO- or FGFRi-treated samples (P < 0.001, P < 0.0001; Fig. 3C–E and Additional file 4: Fig. S4B). Similarly, PIWIL2 was expressed at similar levels in germ cells of DMSO- and FGFRi-treated samples but was variable in MEKi-treated samples, with some germ cells staining strongly for PIWIL2 and others negative (Fig. 3F and Additional file 4: Fig. S4C). This was not possible to confirm using flow cytometry because a reliable PIWIL2 flow assay could not be developed.
MEK1/2 inhibition increased STRA8, but failed to properly induce meiosis in XY germ cellsSince MEKi inhibited mitotic arrest and male germline differentiation, the expression of female germline markers was investigated to determine if FGFRi or MEKi induced female development in XY germ cells. As expected, the pre-meiotic marker STRA8 was detected in the germ cells of XX E12.5 + 72 h DMSO-treated gonads but was not detected in XY DMSO controls (Fig. 4A and Additional file 5: Fig. S5A). While some germ cells appeared very weakly positive for STRA8 in FGFRi-treated samples, STRA8-positive germ cells were commonly found in MEKi-treated gonads, particularly in germ cells close to the mesonephric-gonadal boundary (Fig. 4A and Additional file 5: Fig. S5A). However, while STRA8 staining was localised in the germ cell nucleus in XX controls, it was detected in the germ cell cytoplasm and nucleus in MEKi treatments, indicating that nuclear import–export also regulates STRA8 activity (Fig. 4A and Additional file 5: Fig. S5A). Flow cytometry demonstrated that 74% of germ cells were STRA8 positive in XX DMSO controls, but only 5% and 7% were STRA8 positive in XY DMSO control and FGFRi-treated samples (Fig. 4B). The proportion of STRA8-positive germ cells in MEKi-treated gonads was 43%, significantly higher than XY controls (P < 0.0001), but lower than XX controls (P < 0.0001, Fig. 4B).
Fig. 4
MEK1/2 signalling inhibition permitted STRA8 expression but failed to effectively induce meiosis in XY germ cells. Analysis of XY or XX E12.5 gonad-mesonephros tissue cultured with DMSO or 500 nM FGFRi or MEKi for 72 h (A–C) or 96 h (D–F). A Immunofluorescent images demonstrating STRA8 localisation. Top panel: DAPI (blue), MVH (green), STRA8 (red), SMA (cyan). Bottom panel: STRA8 (grey). B Percentage STRA8 positive germ cells determined by flow cytometry. C,D Immunofluorescent images demonstrating SYCP3 (C) and phospho-γH2AX (p-γH2AX) localisation. Left panel: DAPI (blue), MVH (green), SYCP3 (red; C) or SMA (red; D) and p-γH2AX (cyan). Middle panel: SCP3 (grey; C). Right panel: p-γH2AX (grey). E Percentage p-γH2AX-positive germ cells determined by flow cytometry. F Flow cytometric cell cycle analysis of G0/G1, S-phase and G2/M based on the incorporation of EdU (S-phase) and propidium iodide (DNA content). A, C, D scale bar: 50 μm. Replicates: A, C, D n = 3–4, B n = 8, E,F n = 8–10. Statistics: B Ordinary one-way ANOVA with Tukey’s multiple comparison, E Brown-Forsythe and Welch ANOVA with Dunnett’s T3 multiple comparisons, F Repeated measures two-way ANOVA with Tukey’s multiple comparisons. Error bars: mean ± SEM. Significance between controls and treatment: *P < 0.05, **P < 0.01, ****P < 0.0001
To determine whether germ cells in FGFRi- or MEKi-treated XY gonads had entered meiosis, gonad sections were triple stained using antibodies specific for SYCP3 (synaptonemal complex protein 3), phosphorylated γH2AX (p-γH2AX) and MVH (Fig. 4C and Additional file 5: Fig. S5B). SYCP3 and p-γH2AX were detected in most germ cells in 72 h XX DMSO controls but not in XY DMSO- or FGFRi-treated gonads (Fig. 4C and Additional file 5: Fig. S5B). A small number of germ cells were positive for SYCP3 in MEKi-treated gonads and a subset also stained for p-γH2AX (Fig. 4C and Additional file 5: Fig. S5B). In addition, rare cells positive for p-γH2AX were detected in XY control, FGFRi- and MEKi-treated gonads (Additional file 5: Fig. S5B), however, most did not express MVH and were likely to be apoptotic somatic cells in which p-γH2AX also marks double strand DNA breaks.
To test the possibility that meiotic entry of germ cells in FGFRi- or MEKi-treated gonads was delayed, we cultured E12.5 XX and XY gonad-mesonephros samples for 96 h with DMSO or 500 nM FGFRi or MEKi. IF staining revealed that most XX DMSO germ cells were p-γH2AX positive, indicating they had entered meiosis (Fig. 4D and Additional file 5: Fig. S5C). p-γH2AX-positive germ cells were rarely detected in XY DMSO- or FGFRi-treated gonads but were more common in MEKi-treated samples (Fig. 4D and Additional file 5: Fig. S5C). Quantification using flow cytometry revealed that 88% of germ cells were p-γH2AX positive in XX DMSO samples while only 2%, 4% and 21% were p-γH2AX positive in XY control, FGFRi- and MEKi-treated samples, respectively (Fig. 4E).
Cell cycle analysis of germ cells from the same gonads using EdU (S-phase) to quantify DNA synthesis and PI to measure DNA content demonstrated that the majority of germ cells in E12.5 + 96 h cultures were in G2/M in XX DMSO controls, but were in G0/G1 in XY DMSO- and FGFRi-treated gonads (Fig. 4F). Significantly more germ cells were in G2/M in MEKi than in XY DMSO control (P < 0.05) or FGFRi cultures (P < 0.01), but remained less than in XX controls (P < 0.0001, Fig. 4F). Therefore, while MEKi treatment resulted in a significantly greater percentage of p-γH2AX expressing germ cells that were in G2/M, this proportion was substantially lower than in XX controls indicating that meiosis was not properly induced within the normal temporal window following MEK1/2 inhibition.
The majority of transcriptional divergence occurs after E12.5 in XY and XX germ cellsWe next used RNA sequencing to gain greater insight into genome-wide transcriptional changes in fluorescent activated cell sorting (FACS) isolated Oct4-eGFP-positive germ cells of E12.5 XX and XY gonads (Time 0 controls) and gonads cultured for 24 and 72 h with DMSO, FGFRi and MEKi (Fig. 5A). Differential expression analysis identified 183 and 234 genes that were expressed higher in XY and XX germ cells at E12.5, respectively (time 0; false discovery rate (FDR) < 0.05; absolute fold-change (FC) ≥ 1.5, absolute logFC ≥ 0.585; Fig. 5B, Additional file 6: Table S1.1–1.2). Included in the differentially expressed genes (DEGs) that were higher in E12.5 XY germ cells were a range of Nodal signalling associated genes, including Nodal, Tdgf1 (Cripto), Lefty1, Lefty2, Pitx2 and Otx2, which are known to be high in XY germ cells at this time point [38, 41,42,43] (Additional file 6: Table S1.1). E12.5 XX germ cells expressed higher levels of BMP target genes, including Msx1, Msx2, Id1, Id2, Id3, Stra8 and Gata2, consistent with observations that BMP2 promotes female germline development [43, 44] (Additional file 6: Table S1.2). However, despite these sex-specific transcriptional differences in E12.5 germ cells, our functional data strongly indicated that these differences were insufficient to ensure male germline commitment as MEK1/2 inhibition at E12.5 substantially disrupted male germline differentiation (Figs. 1 and 3).
Fig. 5
FGF-MEK1/2 signalling supports expression of stem cell-associated genes in early germ cells, but only MEK1/2 signalling is required for male germline differentiation. RNA sequencing analysis of germ cells from XX or XY E12.5 gonads, or XX or XY E12.5 gonads cultured for 24 or 72 h with DMSO or 500 nM FGFRi or MEKi. A Example of FACS scatterplot depicting GFP-positive germ cell isolation. B Number of differentially expressed genes (DEGs) between XX or XY E12.5 (time 0) and XX or XY DMSO controls from 24 and 72 h cultures. C Multidimensional scaling (MDS) of all control conditions. D MDS of XX and XY gonads cultured for 24 h. E Differential gene expression analysis of XY E12.5 + 24 h FGFRi XY (i) or XY E12.5 + 24 h MEKi (ii) vs XY E12.5 + 24 h DMSO. F Venn diagram of 24 h FGFRi and MEKi DEGs (i) and heatmap of common DEGs (ii). Asterisks represent genes associated with germ cell tumours, cancer/stem cell biology and/or proliferation. G MDS of XX and XY gonads cultured for 72 h. H Differential gene expression analysis of XY E12.5 + 72 h FGFRi (i) or XY E12.5 + 72 h MEKi (ii) vs XY E12.5 + 72 h DMSO. I Venn diagram comparing MEKi 72 h culture DEGs expressed lower (i) or higher (ii) than expected with XX or XY specific genes identified in B. J Heatmap of DEGs identified in XY E12.5 + 72 h MEKi vs XY E12.5 + 72 h DMSO associated with male germline differentiation, DNA methylation/piRNA pathway, cell cycle, female germline differentiation/meiosis, Nodal signalling, pluripotency and spermatogonial stem cells (SSCs). Asterisks highlight genes associated with cell cycle, meiosis and pluripotency but are not differentially expressed. For all comparisons, genes with FDR < 0.05 and |logFC|> 0.585 (equivalent to |FC|> 1.5) were considered differentially expressed
To identify male and female transcriptional changes that occurred as a normal part of sex-specific germline differentiation, XX and XY germ cells of DMSO control samples were compared after 24 and 72 h of culture. Multidimensional scaling (MDS) revealed that although time 0 XY and XX E12.5 samples were different, they diverged substantially more after 24 and 72 h of gonad culture (Fig. 5C). Differential gene expression analysis revealed that 211 and 3437 genes were increased, and 217 and 2160 genes were decreased in XY germ cells after 24 and 72 h compared to E12.5 (time 0) XY germ cells, respectively (Fig. 5B, Additional file 6: Table S1.3–1.4). Similarly, 340 and 3214 genes were increased, and 157 and 2592 genes were decreased in XX germ cells compared to E12.5 XX germ cells after 24 and 72 h (Fig. 5B, Additional file 6: Table S1.5–1.6). Together, these data indicated that while male and female germline differentiation progressed in the first 24 h, the greatest transcriptional change occurred between 24 (~ E13.5) and 72 h (~ E15.5).
To identify genes specifically associated with male and female germline differentiation, we compared XY with XX germ cells from DMSO controls. This revealed 589 and 3548 genes higher in XY than XX germ cells after 24 and 72 h (XY germline genes; Fig. 5B, Additional f
Comments (0)