
記住我
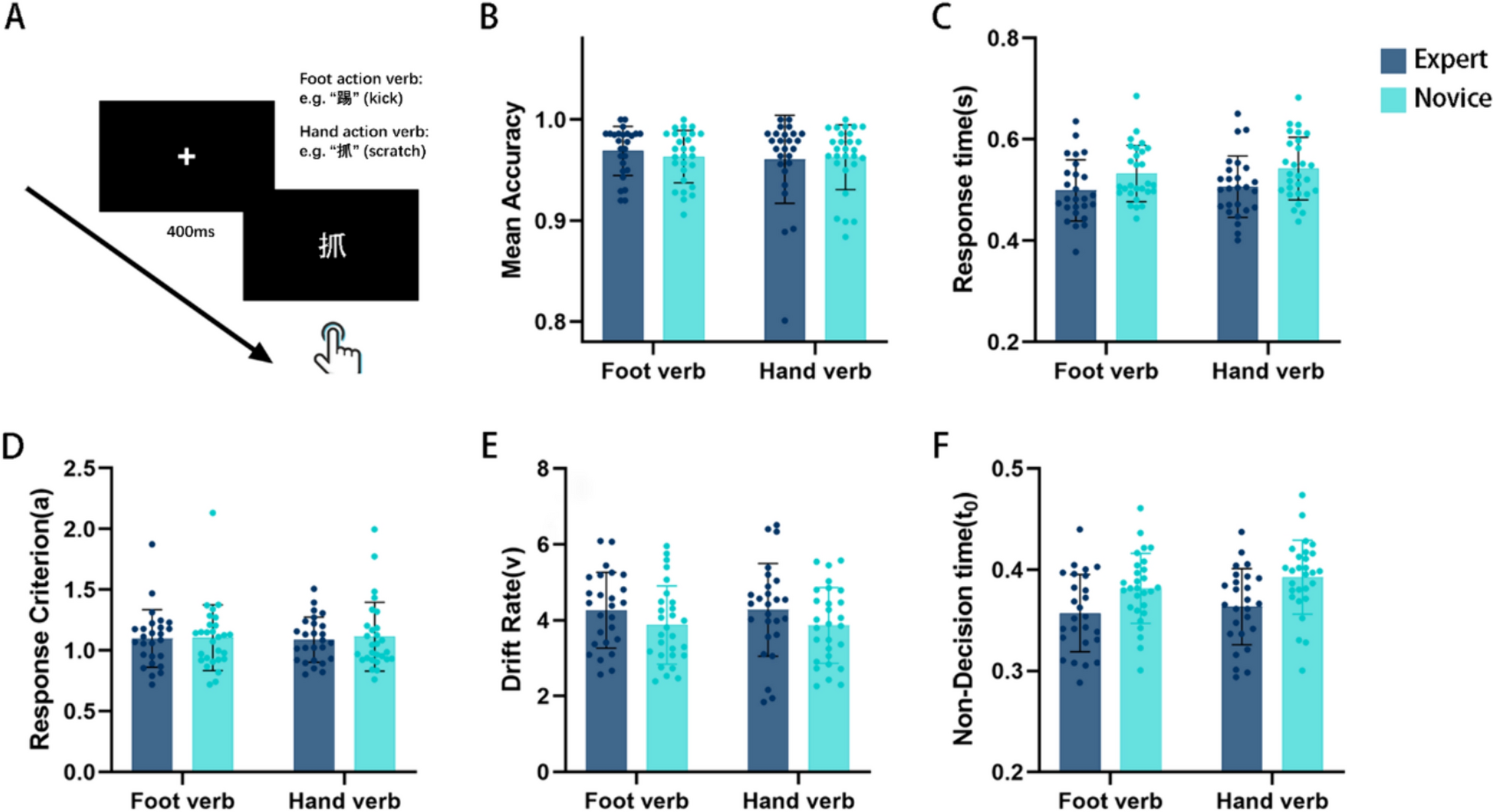





Medial rectus motoneurons showed a phasic-tonic firing profile that was found linearly correlated to eye position and eye velocity. During fixations, they increased their firing rate for eye positions located towards the on direction, which was contralateral to the recording side (i.e., the nasal direction of the ipsilateral eye). Since recordings were carried out in the right oculomotor nucleus, this means that discharge frequency was higher when gaze was directed toward the left side (Fig. 2A). In addition, medial rectus motoneurons displayed a phasic component which was present for rapid eye movements. They exhibited a brisk burst of spikes for saccades in the on direction (Fig. 2A, solid dots) and an abrupt decay in firing or a pause for saccades in the off direction (Fig. 2A, asterisks). ABD Ints exhibited a tonic-phasic discharge pattern that was similar to that of medial rectus motoneurons, except that their on direction was the opposite, i.e., towards the ipsilateral side of the recording (i.e., the temporal direction of the ipsilateral eye). As recordings were obtained in the left abducens nucleus, their discharge increased for eye movements towards the left side. Nevertheless, ABD Ints carry appropriate information to medial rectus motoneurons, since their axons cross the midline and terminate via excitatory synapses on these motoneurons located in the contralateral oculomotor nucleus (Fig. 1; Highstein and Baker 1978). Thus, given that medial rectus motoneurons were recorded in the right oculomotor nucleus and ABD Ints in the left abducens nucleus, the on-direction of both cell types was the same, i.e., toward the left (Fig. 2B). In general, it was observed that ABD Ints fired at higher tonic-phasic rates than medial rectus motoneurons, in particular, the firing bursts for on-directed saccades reached higher frequencies (Fig. 2B, solid dots). Similar to medial rectus motoneurons, ABD Ints rapidly decreased or stopped firing during off-directed saccades (Fig. 2B, asterisks).
Fig. 2
Firing of MR motoneurons, ABD Ints, and ATD neurons, during spontaneous eye movements. The figure shows examples of the firing rate (FR, in spikes/s) during spontaneous eye movements of the three neuronal types studied: A MR motoneurons (Mns), B internuclear neurons (Ints) of the ABD, and C vestibular neurons of the ATD. L and R next to the double arrow in A stands for leftward and rightward eye movements, respectively (for A–C). MR motoneurons and ATD neurons were recorded in the right side, and ABD Ints in the left side. The horizontal eye position (EP, eye position, in degrees) and velocity (EV, eye velocity, in degrees/s) are shown for the eye ipsilateral to the recording site in (A–C). Note that the discharge of the three neurons increases with eye movements toward the left side, although the correlation between FR and EP and EV can be better appreciated in the ABD Int (B) and the MR Mn (A) as compared to the ATD neuron (C). Solid dots point to burst-like increases in FR during on-directed saccades, asterisks indicate an abrupt decay in FR for off-directed saccades, and arrowheads show an example of absence of FR response during saccades in the ATD neuron (C)
During spontaneous eye movements, the discharge of ATD neurons differed somewhat from that of motoneurons and ABD Ints. Thus, as can be appreciated in Fig. 2C, although they also modulated in relation to eye movements, their degree of correlation with eye position was less conspicuous. They increased their discharge rate as eye positions were fixed more to the contralateral side (left) of the recording (right) site, as medial rectus motoneurons did. Since ATD neurons send excitatory projections to the ipsilateral oculomotor nucleus (Baker and Highstein 1978), the signals conveyed by these premotor cells were appropriate for driving medial rectus motoneurons. Therefore, the three neuronal types recorded as illustrated in Fig. 1 presented eye movements to the left as the on-direction. The behavior of ATD neurons also showed a phasic component. During on-directed saccades, they discharged a burst of spikes, which was, however, of low frequency (Fig. 2C, solid dots), and on occasions, it was not present (Fig. 2C, arrowheads). For off-saccades, ATD neurons usually decreased their firing rate (Fig. 2C, asterisks), and rarely they showed no change in their discharge activity or this was not consistent. Thus, their phasic activity was present (Fig. 2C), but it was not as precise as that of medial rectus motoneurons (Fig. 2A) and ABD Ints (Fig. 2B).
Quantitative comparison of the signals displayed during spontaneous eye movements in ATD neurons, ABD Ints and their target medial rectus motoneuronsA comparison of eye-related parameters between the three neuronal types was carried out during spontaneous eye movements. Although our principal aim was the comparison between ATD neurons and ABD Ints, we also included the discharge properties of medial rectus motoneurons to discern the degree of similarity in the firing activity of both inputs with respect to their target motoneurons. The number of neurons analyzed during spontaneous eye movements was n = 25 medial rectus motoneurons, n = 14 ABD Ints, and n = 13 ATD neurons.
During fixations, neuronal eye position sensitivity (ks) was calculated as the slope of the regression line between firing rate and eye position. The result of this analysis for the three neuronal populations illustrated in Fig. 2 is shown in Fig. 3A. It can be appreciated that the slopes, and therefore ks values, of the ABD Int and the medial rectus motoneuron, were higher than that of the ATD neuron.
Fig. 3
Quantitative comparison of eye-related sensitivities obtained during spontaneous fixations and saccades between MR motoneurons and their two major pontine inputs. A Correlation between firing rate (FR, in spikes/s) and eye position (EP, in degrees) was carried out by linear regression analysis. The slope of the regression line thus obtained represents the neuronal eye position sensitivity (ks, in spikes/s/degree). The three lines correspond to the neurons shown in Fig. 2, whose ks values were 4.1, 7.9, and 2.1 spikes/s/degree, for the MR Mn of Fig. 2A, the ABD Int of Fig. 2B, and the ATD neuron of Fig. 2C, respectively. B Box-and-whisker plot showing eye position sensitivity during eye fixations (ks, in spikes/s/degree) between MR Mns, ABD Ints, and ATD neurons. One-way ANOVA test revealed significant differences (p ≤ 0.001) between groups. Pairwise multiple comparisons (Dunn’s method) demonstrated that ABD Ints and MR Mns showed significantly higher ks values than ATD neurons (p < 0.05 for both cases; asterisks). C Same as B but for the eye position threshold at which the neuron was recruited into activity (Th, in degrees). One-way ANOVA test revealed significant differences (p ≤ 0.001) between groups. All pairwise multiple comparisons (Dunn’s method) were significantly different (p < 0.05 for the three cases; asterisks). ATD neurons showed the lowest threshold and MR Mns the highest threshold. D Same as B but for eye velocity sensitivity during saccades (rs, in spikes/s/degree/s). Significant differences were present between the three groups (one-way ANOVA, p ≤ 0.001). When pairwise multiple comparisons were carried out (Dunn’s method) all of them resulted in significant differences (p < 0.05). ABD Ints showed the highest rs values, while ATD neurons presented the lowest. For B–D, n = 25 MR Mns, 14 ABD Ints, and 13 ATD neurons
When neuronal eye position sensitivities (ks) were compared between the three neuronal populations, it was found that ks values of ATD neurons were significantly lower than those of both ABD Ints and medial rectus motoneurons (Fig. 3B; one-way ANOVA on ranks followed by Dunn’s method for post hoc comparisons, p ≤ 0.001, H = 31.470, Cohen’s d = 2.287). Mean ± SEM ks values were 4.81 ± 0.24, 6.62 ± 0.57, and 1.81 ± 0.3 spikes/s/degree, for medial rectus motoneurons, ABD Ints, and ATD neurons, respectively. Moreover, ks values of ABD Ints were similar to those of motoneurons (p > 0.05) whereas those of ATD neurons were significantly lower (p < 0.05) (Fig. 3B). It should be noted that, however, the statistical comparison of the coefficients of determination (R2) for the rate-position regression equations between the three populations showed no statistical difference (ANOVA on ranks, p = 0.158, H = 3.696), indicating that the ks signal was present in the two premotor inputs, as well as in the motoneurons.
The eye position threshold for recruitment into activity was also calculated from the regression equations and compared between the three groups. Interestingly, in this case, there was a significant difference between all neuronal populations, so that ATD neurons showed the lowest threshold and medial rectus motoneurons the highest (Fig. 3C; one-way ANOVA on ranks followed by Dunn’s multiple comparisons, p ≤ 0.001, H = 28.013, d = 2.001). Therefore, the recruitment threshold of the two afferent populations (ATD neurons and ABD Ints) was lower than that of the motoneurons they terminate on. In turn, the threshold of ATD neurons was significantly (p < 0.05) lower than that of ABD Ints, that is, they were recruited into activity at more negative (off-directed) eye positions in the orbit. We would like to highlight that the results in threshold were quite striking, particularly due to the extremely low threshold of ATD neurons as compared to ABD Ints and medial rectus motoneurons (mean ± SEM were: − 36.37 ± 5.11 degrees, − 13.71 ± 1.46, and − 8.17 ± 0.54, respectively).
During saccades, the neuronal eye velocity sensitivities (rs) obtained for the three populations reflected the qualitative observations described above from their respective firing pattern. Thus, ABD Ints showed the highest rs values (1.26 ± 0.06 spikes/s/degree/s), and ATD neurons had the lowest rs values (0.19 ± 0.04 spikes/s/degree/s). Medial rectus motoneuronal data were in between (0.50 ± 0.02 spikes/s/degree/s). The differences between the three groups reached statistical significance (Fig. 3D; one-way ANOVA on ranks followed by Dunn’s multiple comparisons, p ≤ 0.001, H = 42.109, d = 3.673). The saccadic signal of ATD neurons was weak and not always consistent. This yielded low correlations of determination of the rate-velocity regression equations for ATD neurons, which were significantly lower than those obtained for both ABD Ints and motoneurons (one-way ANOVA followed by Holm–Sidak’s multiple comparisons, F(2,49) = 60.517, p ≤ 0.001). It should be highlighted that with respect to all eye-related parameters calculated in the present work, only eye velocity sensitivity for spontaneous saccades (rs) produced significantly lower coefficients of determination in ATD neurons compared to ABD Ints and medial rectus motoneurons. In all other parameters, the three groups of neurons showed similar coefficients of determination. Therefore, the saccadic signal in ATD neurons was not as accurate as that of the other main input to the medial rectus motoneurons, the ABD Ints.
Discharge pattern of medial rectus motoneurons, ABD Ints, and ATD neurons during vestibular eye movementsDuring vestibularly induced eye movements the firing profile of medial rectus motoneurons and that of ABD Ints was similar (Fig. 4A, B). The two neuronal types modulated during both the slow and the fast phases of the nystagmus. However, whereas the discharge of medial rectus motoneurons increased for head rotations towards the same side of the recording (type I response; Gernandt 1949), ABD Ints increased their firing for head rotations towards the opposite side (type II response; Gernandt 1949). As mentioned above, as ABD Ints were recorded in the left side and medial rectus motoneurons in the right side, then both neuronal types increased their discharge for rightward head rotations resulting in eye movements to the left. As can be observed in Fig. 4A, B, although they showed the same discharge pattern, ABD Ints reached, in most cases, higher firing frequencies than medial rectus motoneurons during vestibular eye movements.
Fig. 4
Behavior of MR Mns, ABD Ints, and ATD neurons during vestibular eye movements. Traces in A–C illustrate from top to bottom: eye position (EP, in degrees), eye velocity (EV, in degrees/s), and firing rate (FR, in spikes/s). The red trace in EV represents head velocity, which is shown inverted for clarity. L and R next to the double arrow in A indicate leftward and rightward eye movements, respectively (for A–C). MR motoneurons and ATD neurons were recorded in the right side, and ABD Ints in the left side. Note that in the three neuronal types (A–C), on-directed eye movements were those toward the left. The firing pattern of MR Mns (A) and ABD Ints (B) during vestibularly induced eye movements was similar. Thus, their discharge increased for slow and fast phases of the nystagmus in the on direction and decreased for slow and fast phases in the opposite direction. However, ATD neurons clearly modulated during the slow phases of the vestibulo-ocular reflex but lacked any signal related to fast phases (C). Note also that whereas ATD neurons and MR Mns showed type I response during vestibular eye movements (i.e., their discharge increased for head rotations toward the ipsilateral side of the recording), ABD Ints showed type II response (i.e., firing increased for head rotations toward the contralateral recording side)
The response of ATD neurons during vestibularly induced eye movements was type I since their firing increased for head rotations towards the ipsilateral side of the recording (Gernandt 1949; Markham et al. 1986). During vestibular eye movements, their firing pattern differed from that of motoneurons and ABD Ints (Fig. 4C). They modulated sinusoidally (as the stimulus) without showing bursts or decreases in activity for the on- and off-directed fast phases of the nystagmus, respectively, in marked contrast to motoneurons and ABD Ints (Fig. 4A, B). This behavior likely corresponds to the head velocity signal described by Reisine and Highstein (1979).
Quantitative comparison of ATD neurons and ABD Ints signals during vestibular eye movements in relation to medial rectus motoneuronsEye position and eye velocity sensitivities were calculated during vestibular eye movements (namely kv and rv, respectively) selecting the slow phases of the nystagmus and using multiple regression analysis. ABD Ints showed higher kv values than ATD neurons (Fig. 5A; one-way ANOVA on ranks followed by Dunn’s multiple comparisons, p ≤ 0.001, H = 25.816, d = 1.838). Mean ± SEM data were 7.18 ± 0.63 and 0.83 ± 0.18 spikes/s/degree, respectively. When compared with medial rectus motoneurons (5.31 ± 0.37 spikes/s/degree), ABD Ints showed similar kv (p > 0.05), whereas kv in ATD neurons was significantly lower than in the motoneurons (p < 0.05) (Fig. 5A).
Fig. 5
Eye-related parameters of MR Mns, ABD Ints, and ATD neurons during vestibular eye movements. A Box-and-whisker plot illustrating the values of neuronal eye position sensitivity during the slow phases of the vestibular nystagmus (kv, in spikes/s/degree) for MR Mns, ABD Ints, and ATD neurons. One-way ANOVA on ranks detected significant differences between groups (p ≤ 0.001). Post-hoc pairwise multiple comparisons (Dunn’s method) revealed that MR Mns and ABD Ints showed significantly (p < 0.05, asterisks) higher kv values than ATD neurons. B Box-and-whisker plot showing the values of neuronal eye velocity sensitivity during the slow phases of the vestibulo-ocular reflex rv (in spikes/s/degree/s). One-way ANOVA on ranks revealed that there were significant differences between groups (p ≤ 0.001). When pairwise multiple comparisons were performed (Dunn’s method), we obtained that ABD Ints showed significantly higher rv values than both ATD neurons (p < 0.05) and MR Mns (p < 0.05). The number of neurons analyzed in A and B was 25 MR Mns, 13 ABD Ints, and 9 ATD neurons
Regarding eye velocity sensitivity (rv), ABD Ints also exhibited higher values (1.37 ± 0.06 spikes/s/degree/s) than ATD neurons (0.87 ± 0.08 spikes/s/degree/s) (Fig. 5B; one-way ANOVA on ranks followed by Dunn’s method for post hoc comparisons, p ≤ 0.001, H = 26.054, d = 1.856). However, in this case, eye velocity sensitivity of ATD neurons during vestibular eye movements (rv) was similar (p > 0.05) to that of motoneurons (0.71 ± 0.05 spikes/s/degree/s), whereas ABD Ints showed significantly higher rv values than motoneurons and ATD neurons (p < 0.05) (Fig. 5B).
留言 (0)