Schistosomiasis is the second most serious transmissible disease owing to its great impact on infected subject morbidity and mortality (Colley et al. 2017). The main complication of schistosomiasis as variceal bleeding secondary to liver fibrosis and portal hypertension is crippling and yet, life threatening. Therefore, prevention of fibrosis remains the only feasible strategy available (Kong et al. 2020). Though very few agents (e.g., pirfenidone and nintedanib) have recently been approved for pulmonary fibrosis, an effective antifibrotic drug that can be used safely in liver fibrosis is considered a big challenge (Fang et al. 2021). Therefore, promising plant-derived compounds possessing pleiotropic pharmacological actions with a long history of safety, such as resveratrol, merits attention (Koushki et al. 2018).
In this study, we document an evident antifibrotic effect of resveratrol in schistosomal hepatic fibrosis and delineate some contributing mechanisms. In accordance with previous studies (Fang et al. 2021) (El-Lakkany et al. 2012), the untreated S. mansoni infected mice showed an elevation in serum levels of liver enzymes reflecting the loss of hepatocyte integrity, whereas the detected hypoalbuminemia could be the result of decreased albumin synthesis from damaged hepatocytes or due to decreased absorption from damaged intestinal mucosa (Al Humayed et al. 2020; El-Lakkany et al. 2012).
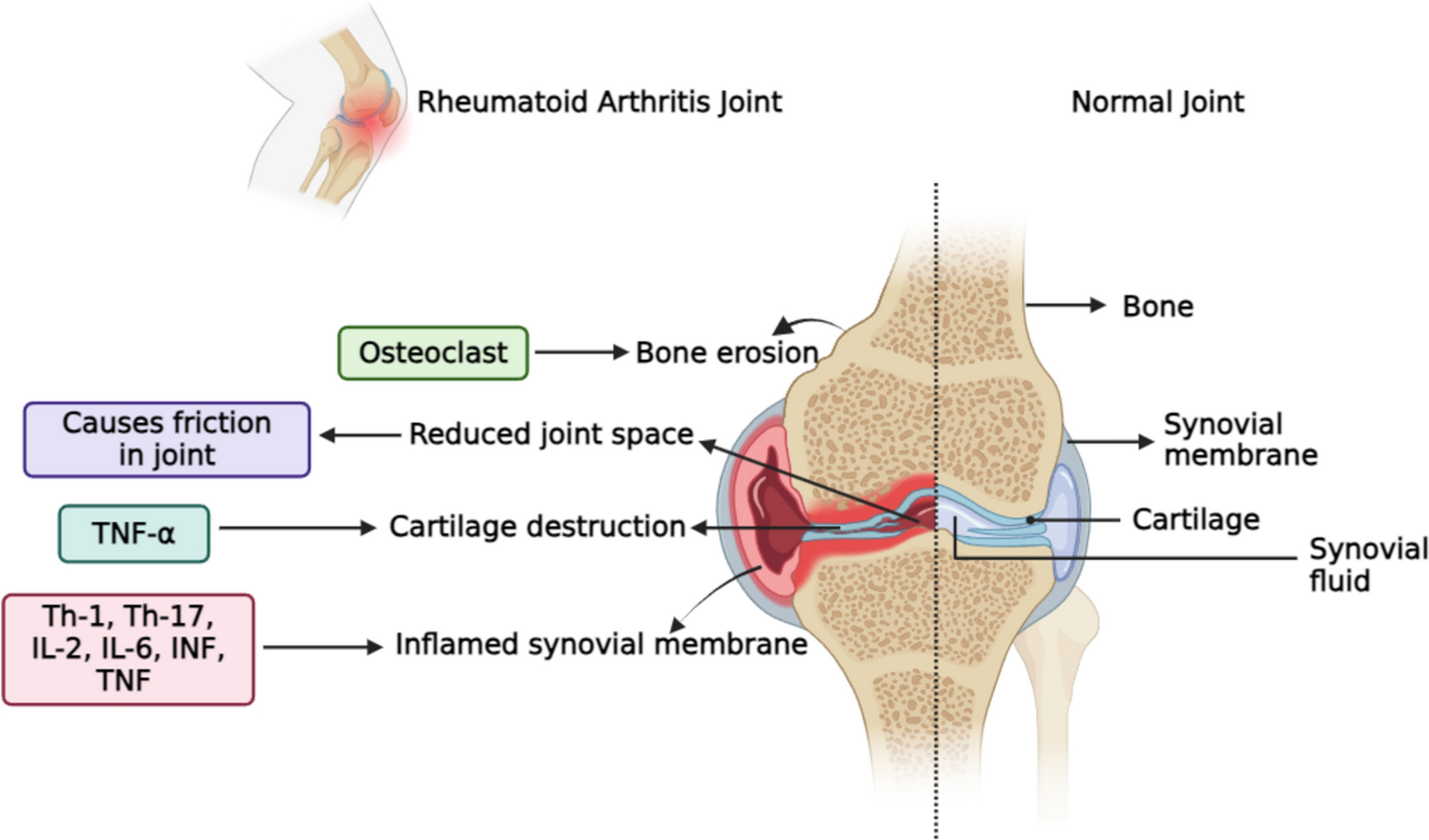
Hepatocyte damage occurs secondary to the depicted massive hepatic proinflammatory state evidenced by the significant elevated expression of the inflammatory cytokines and chemokine genes with subsequent stimulation of the fibrotic process. In fact, HSCs are recognized to serve as the primary target for many inflammatory stimuli of fibrogenesis (Hou and Syn 2018; Seki and Schwabe 2015). It is thus expected that suppression of the release of inflammatory mediators from injured hepatocytes and Kupffer cells is crucial for mitigating HSCs activation and consequently suppression of fibrogenesis (El-Agamy et al. 2011).
Additionally, the observed upregulation of TGF-β1, α-SMA and ultimately collagen 1 coincides with the demonstrated histological findings of S. mansoni-induced liver injury. TGF-β1 is a pleiotropic cytokine that displays proinflammatory, fibrogenesis, and immunosuppressive properties (Mantel and Schmidt-Weber 2011). In the context of liver schistosomiasis, TGF-β1 expression has been demonstrated to be increased in liver tissues from animal models as well as patients with liver schistosomiasis (Chen et al. 2008; El-Lakkany et al. 2011, 2012; Li et al. 2014a, b). It plays a role in activation and differentiation of HSCs into myofibroblast phenotype that expresses α-SMA and deposits excessive amount of extracellular matrix, predominantly collagen type I (Gressner and Weiskirchen 2006). In turn, the activated HSCs stimulate TGF-β, and a vicious circle ensues (Wahl et al. 1997). The critical role of TGF-β1 was further emphasized by the ability of TGF-β1 soluble receptors (Yata et al. 2002) or TGF-β neutralizing antibody to scavenge TGF-β1 and reverse hepatic fibrosis in experimental animals (Ling et al. 2013). Nevertheless, a substantial body of evidence highlights other pathways contributing to schistosomal fibrosis which either minimally involves TGF-β1 or is completely TGF-β1 independent (Kaviratne et al. 2004; Liu et al. 2014).
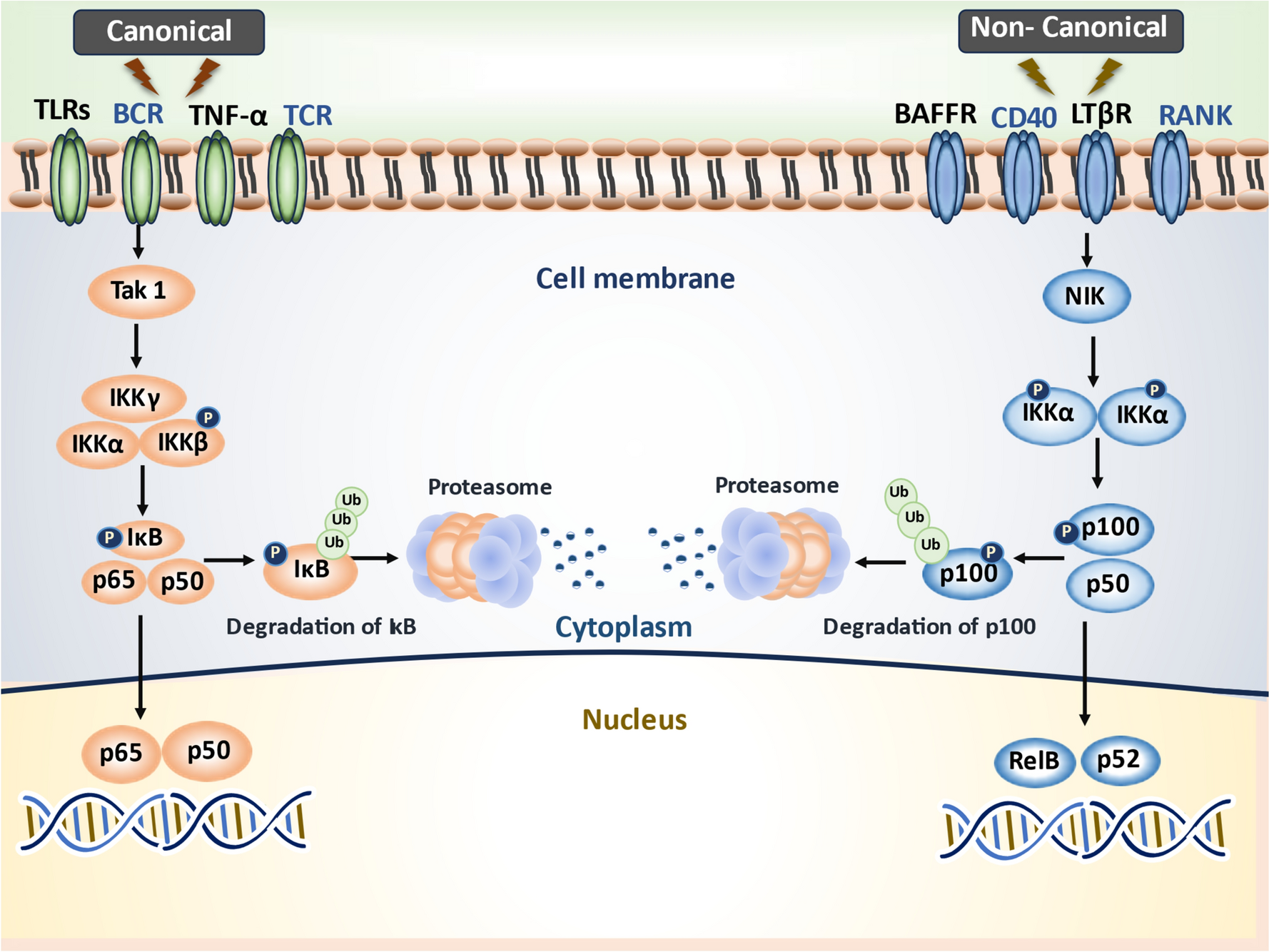
Herein, we detected a significant increase in phosphorylation of the Iκβ which triggers its recognition by an E3 ligase, its polyubiquitination and subsequent degradation by the proteasome. Such degradation will release and thus switch on the translocation of NF-κB, the master key of the inflammatory response to the nucleus activating cytokines, chemokines, and immunoreceptors gene expression in addition to the profibrogenic cytokine TGF-β. The functional nature of NF-κB target genes underpins the harmful consequences of its dysregulated activity and necessitates its tight control (Christian et al. 2016). Accumulating data points to the essential role of NF-κB in promoting HSCs activation and survival, and engendering fibrosis (Marcher et al. 2019; Muriel 2009). In this context, an increase in NF-κB binding activity in liver tissues from Schistosoma japonicum infected mice was previously reported confirming its role in mediating hepatic schistosomiasis (Chen et al. 2008).
The complex interaction of the different pathways contributing to hepatic fibrosis would finally lead to portal hypertension associated with splanchnic vasodilation (Abraldes et al. 2006). Consistent with previous studies reporting the development of a state of hyperdynamic circulation in human as well as animal models of portal hypertension (Bolognesi et al. 2014; Sarin et al. 1991; Strauss 2002), we detected a significantly elevated portal pressure associated with a significant increase in the mesenteric blood flow when measured 10 weeks post cercarial exposure. Clinically wise, splenomegaly, as detected here, is an important manifestation of abnormally raised portal pressure (Kong et al. 2020). It has been suggested that splenomegaly plays a role in the progression of liver fibrosis to cirrhosis. Although the precise mechanism has not been fully elucidated, the excessive production of TGF-β1 from splenic macrophages could be a critical contributing factor (Li et al. 2017).
Although resveratrol has gained much interest in the management of liver diseases (Bishayee et al. 2010), its effect on portal pressure in S. mansoni-infected mice was not addressed in previous works. Our results showed a significant decrease in portal pressure, portal flow, spleen weight by both the low and high doses of resveratrol.
The pathophysiologic mechanisms of portal hypertension involve increased flow in portal vessels and/or increased intrahepatic vascular resistance (Gunarathne et al. 2020), The former occurs as a consequence of splanchnic vasodilation whereas the latter reflects hepatic architectural changes involving myofibroblasts, HSCs, and vascular smooth muscle cells (VSMCs) which play a central role in regulating the intrahepatic vascular tone. Moreover, dysfunction of sinusoidal endothelium with decreased nitric oxide (NO) production or function greatly affects the tone of the adjacent VSMCs (Mehta et al. 2014). The net result is increased pressure in the portal circulation due to an increase in blood flow.
Treatment with resveratrol produced a reduction in portal pressure that was more prominent with the higher dose. In line, Di Pascoli et al. (2013), reported a favorable effect of resveratrol on portal pressure in carbon tetrachloride (CCl4)-induced cirrhotic rats. This beneficial effect could be ascribed to inhibition of HSC activation secondary to the observed dose-dependent suppression of TGF-β and α-SMA gene expression; both have strong stimulatory effect on HSCs. This suppression was associated with consequent reduction in the expression of collagen I and preservation of liver architecture. Indeed, the gross appearance of the liver tissue obtained from the resveratrol-treated mice was significantly improved especially with the higher dose. This was reflected microscopically on near total mitigation of fibrotic changes, decreased granulomas, and the distinct resolution of inflammatory infiltrations.
In concordance, favorable effects of resveratrol on various models of liver injury have been demonstrated in both in vitro and in vivo studies (Bishayee et al. 2010; Muriel and Rivera-Espinoza 2008). It exerts protective effect against various hepatotoxicants (Al Humayed et al. 2020; Bingul et al. 2021; Lee et al. 2010). Likewise, resveratrol exhibited a beneficial effect against liver diseases such as chronic biliary obstruction (Ara et al. 2005), and non-alcoholic fatty liver disease (Ara et al. 2005; Li et al. 2014a, b).
A previous study examined the effect of resveratrol, among others, as preventive and therapeutic agents against liver fibrosis induced by Schistosoma mansoni infection. Noticeably, they found that resveratrol can prevent the development of liver fibrosis, but it had no effect on reversal of fibrosis (El-Agamy et al. 2011).
Although there has been tremendous progress over the past decade in understanding the antifibrotic effect of resveratrol, the detailed molecular mechanisms remain elusive (Izzo et al. 2021). For instance, resveratrol is well recognized to exert effective antioxidant effects owing to its capability to promote the synthesis and improve the activity of antioxidant molecules. Nevertheless, it failed to prevent markers of oxidative stress in a model of CCl4-induced hepatic injury (Pan et al. 2017). Hence, it is assumed that the anti-fibrotic effect of resveratrol is independent of the sole antioxidant effect.
Several studies reported that natural compounds including resveratrol mitigates various pathological conditions via interfering with NF-κB signaling pathway (Laurindo et al. 2023; Ren et al. 2013; Xu et al. 2018). As formerly discussed, we detected a significant increase in the p-IκB/IκB ratio in the untreated-infected mice which triggers the subsequent release of NF-κB and the enhanced transcription of its target genes as reflected by the increased cytokines and chemokine. However, to date, the effect of resveratrol on translocation, DNA-binding, and phosphorylation of the key components in NF-κB signaling pathway is controversial (Ren et al. 2013; Tsai et al.1999). For instance, Chavez et al. (2008) demonstrated that the antifibrotic effect of resveratrol in chronic CCl4-induced liver was associated with a non-significant decrease in the relative optical density of nuclear NF-κB. It is worth noting that they employed a single dose of resveratrol in their study (10 mg kg/kg orally, for 8 weeks), therefore, a positive effect of a higher dose cannot be ruled out.
On the other hand, an earlier study reported that resveratrol inhibited the nuclear translocation of NF-κB but it showed no effect on the phosphorylation nor degradation of IκBα (Manna et al. 2000). On the contrary, Pellegatta et al. demonstrated that a longer period of incubation with resveratrol could inhibit phosphorylation of NF-κB components and IκBα in human endothelial cells (Pellegatta et al. 2003).
In fact, our finding revealed a dose-dependent inhibition of IκB activity as shown by the significant decrease in the p-IκB/IκB ratio with both doses of resveratrol versus the untreated mice. In line with these data, Ren et al. precluded any effect of resveratrol on DNA-binding activity and transportation of NF-κB. Rather, it blocked the activity, not the expression, of IKBα. Additionally, resveratrol inhibited IκB kinases (IκK) activity that catalyzes the phosphorylation of Iκβ, to further inhibit the canonical activities of NFκB in a dose-dependent manner (Ren et al. 2013).
Remarkably, it has been speculated that resveratrol should act upon upstream molecules to inhibit NF-κB activation by various stimuli. Indeed, data suggest that pharmacological activation of SIRT1 by resveratrol could be a crucial underlying mechanism of its anti-inflammatory, antiproliferative, and antiapoptotic effects (Iside et al. 2020).
Intriguingly, we detected a significantly reduced SIRT1 in the untreated infected mice, which may point to its implication in the pathological consequences of schistosomal infection including fibrosis. Of note, there is a number of critical proofs documenting the protective role of SIRT1 against fibrosis as it controls Smad fibrotic pathway in HSCs. Further, reduced SIRT1 expression or HSC-specific deletion of SIRT1 led to exacerbated fibrosis in mice models (Li et al. 2018; Ren et al. 2022). In line with our data, SIRT1 protein expression was also decreased in liver infected by S. japonicum (Zhou et al. 2021).
SIRT1 has been reported to participate in various physiological and pathological states by controlling expression and activities of several regulatory molecules. Previous studies showed that the protective effect of resveratrol is linked to an increase in SIRT1 activity. For instance, Yu et al. showed that resveratrol reduced the inflammatory response induced by partial liver resection through reduction of high mobility group box 1 (HMGB1) translocation to the nucleus as a result of SIRT1 upregulation (Yu et al. 2019). Furthermore, SIRT1 regulates the expression of antioxidant enzymes, and this effect is implicated for the antioxidant effect of resveratrol (Pan et al. 2017).
Interestingly, an antagonistic crosstalk between NF-κB and SIRT1 signaling pathways in regulating immune and inflammatory response to infection has been uncovered (Kauppinen et al. 2013). This antagonism has double benefits. On one hand, the body needs to respond quickly to harmful stimuli such as an infection or tissue damage by switching to a rapid energy generation system through suppressing SIRT1. On the other hand, it is necessary to switch off the pro-inflammatory milieu once the harmful stimulus has disappeared. The disturbance of this antagonistic balance with sustained stimulation of NF-κB on the expense of active SIRT1 is the centerpiece in chronic liver disease (de Gregorio et al. 2020). Indeed, The HDAC activity of SIRT is the key for its antagonistic effect on NF-kB. Deacetylation of RelA/p65 subunit of NF-κB, impairs its transcriptional activity and further predisposes to its ubiquitination and degradation. The deacetylation of RelA/p65 by SIRT1 also favors the re-association of NF-κB with IκB-α which triggers the back transport of the NF-κB complex from the nucleus to the cytoplasm regaining the inactive state (Rothgiesser et al. 2010; Yang et al. 2010). Thus, the activation of SIRT1 by resveratrol in our study would further explain the suppressed transcription of NF-κB downstream genes, including those encoding IL-1β, TNF-α, TGF- β and MCP-1, and other pro-inflammatory factors (Meng et al. 2021; Tian et al. 2016).
Finally, another debatable point is whether PZQ exerts direct antifibrotic effect aside from its schistosomicidal effect. Previous studies have demonstrated that praziquantel reduced fibrosis as a result of amelioration of the immune and inflammatory responses in different mice models of liver injury (El-Lakkany et al. 2012; Liang et al. 2011; Liu et al. 2014; Singh et al. 2019). Conversely, PZQ was recently reported to have a limited potential for prevention and/or reversal of the already established fibrosis (Nono et al. 2020). In our study, PZQ was administered to both the resveratrol-treated and untreated groups to rule out any significant beneficial effect of praziquantel alone in attenuating fibrosis.
In conclusion, the present study demonstrates that resveratrol could protect against the deleterious hemodynamic and structural consequences of S mansoni-induced liver injury. The beneficial effects of resveratrol can be mainly attributed to its suppressive effect on NF-κB signaling and its downstream proinflammatory and fibrotic transcriptional products. The evidence on the ability of resveratrol to restore the distorted balance of the SIRT1/NF-κB axis, encourages its further clinical evaluation as a promising agent in schistosomal hepatic fibrosis.




Comments (0)