
Remember me
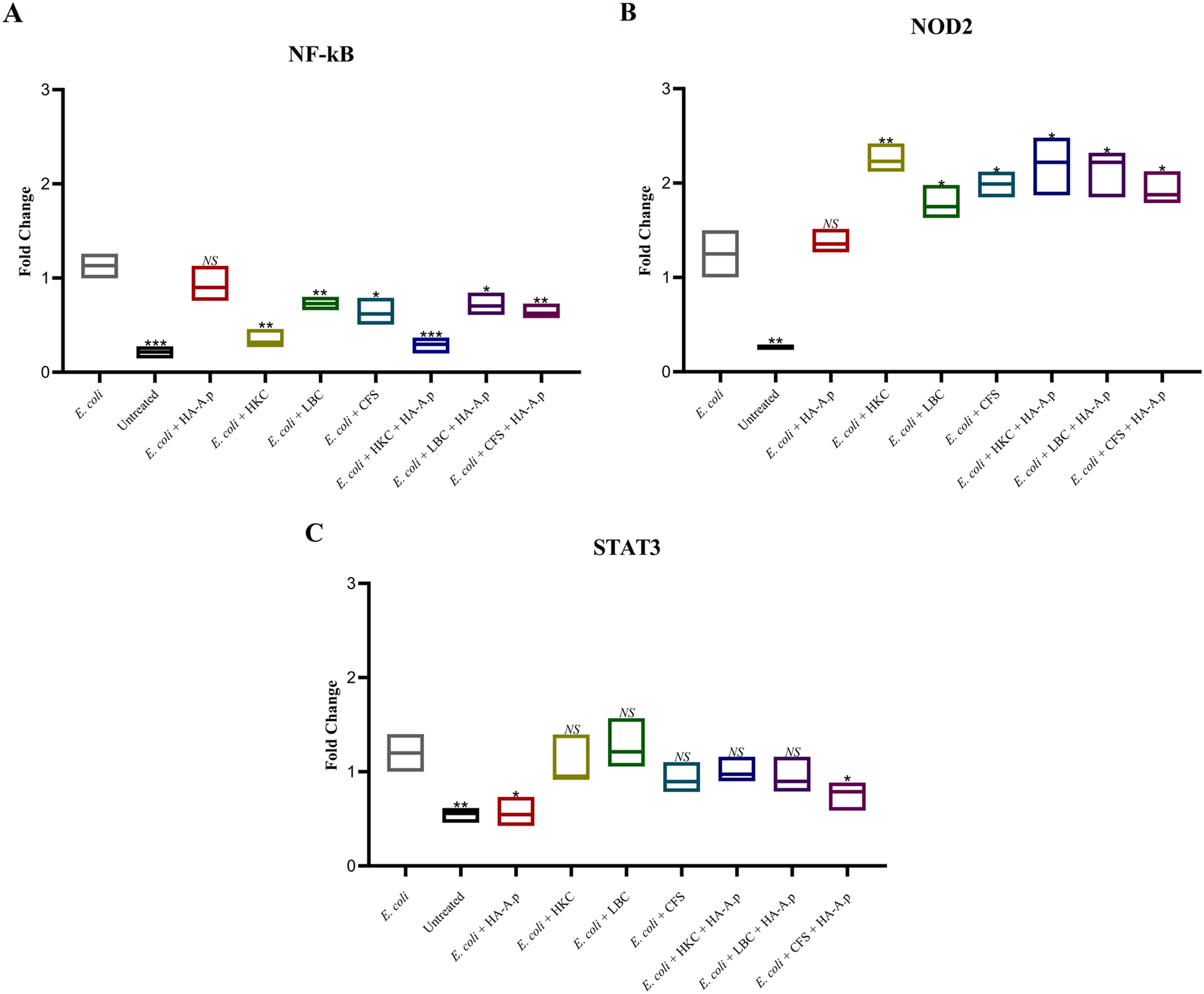


The purpose of rhTNFα exposure is to induce these pro-apoptotic cytokines and observe the apoptotic response, specifically caspase-3 activity in CD34+, CD33+, and CD41 + MDS BMMC. Apoptosis may occur through either the intrinsic or extrinsic pathway. The TNFα apoptosis mechanism model operates through the extrinsic pathway.
The TNFα signaling pathway mechanism for initiating the apoptotic process begins with the binding of TNFα to TNFR-1 and TNFR-2 receptors. TNFR-1 mediates the cell death program by initially binding directly to the adapter protein TNF receptor-associated death domain (TRADD), then indirectly connecting to the FAS-associated protein with a death domain (FADD). This sequential process ultimately leads to the activation of the caspase cascade, culminating in the execution phase facilitated by caspase-3 [16,17,18].
Exposure of rhTNFα to hematopoietic progenitor cells is expected to result in the binding of rhTNFα to its receptor, TNFR1. This binding is anticipated to initiate cell death signals through the death-inducing signaling complex (DISC) and activate TRADD. Subsequently, caspase activation will be triggered, ultimately leading to the activation of caspase-3, which serves as the executor of apoptosis in hematopoietic progenitor cells in MDS, as shown in Fig. 1.
The role of TNFα in triggering apoptosis is supported by a research report that discovered a positive correlation between elevated serum TNFα levels and anemia in MDS patients. Furthermore, an increase in TNFα levels in the bone marrow was linked to heightened apoptosis during the early stages of MDS [19]. Several studies have also documented increased activity of caspase-1 and caspase-3 in hematopoiesis cells of MDS patients, with caspase-3 activity being ten times higher than that of caspase-1. Notably, the heightened caspase-3 activity is positively associated with increased TNFα levels [20, 21].
However, it is possible for caspase-3 activation to occur through interactions aside from rhTNFα exposure, although such instances were negligible in this study. We determined this by calculating the delta value (Δ) for Caspase-3 using the formula: the caspase-3 activity data after rhTNFα exposure subtracted by data before exposure (as the baseline). The positive and statistically significant delta values indicate that rhTNFa exposure can indeed increase caspase-3 activity.
The involvement of TNFα as an apoptosis trigger in MDS was confirmed by the elevated levels of TNFα in the serum of MDS patients. This increase in TNFα showed a positive correlation with clinical anemia, and an elevated rate of apoptosis in the early stages of MDS [19].
In MDS, there is an increasein the activity of both caspase-1 and caspase-3, with caspase-3 activity being ten times higher than caspase-1 [7, 13, 14]. This increased caspase-3 activity is associated with a high level of TNFα [20, 21]. Iriani et al. [22] discovered that the high levels of TNFα in MDS are initiated by an increase in the expression of TNFα mRNA, triggered by sCD40L stimulation.
A substantial portion of MDS cells undergo intramedullary apoptosis, which can be challenging to detect morphologically, as it predominantly occurs at an early stage. Therefore, the assessment of apoptosis in MDS should involve early markers of apoptosis, such as caspase-3.
Apoptosis in CD34+In Fig. 2A, the Δ caspase-3 value in the MDS group was negative, whereas it was positive, with a significant p-value. The Δ value was used to gauge the extent of the impact of rhTNFα exposure. This suggests that, in the MDS group, apoptosis had occurred in CD34 + cells even without exposure to rhTNFα, in contrast to the control group. This implies that CD34 + MDS cells naturally tend to undergo apoptosis.
Reports indicate that CD34 + MDS cells tend to undergo apoptosis, with their susceptibility to apoptosis being most pronounced among CD34 + progenitor cells. [11, 23] This susceptibility is attributed to defects in the progenitor stem cells. Due to these clonal abnormalities, daughter cells would develop into defective cells that are prone to apoptosis. Another study found that MDS CD34 + cells displayed higher apoptotic activity than normal cells, with the apoptosis rate in MDS CD34 + cells being 2.08 times faster than their proliferation rate [24]. In MDS, despite the bone marrow continuing to produce cells, the cell cycle is shorter. It is also known that over two-third of MDS cells at various stages of differentiation in the bone marrow undergo apoptosis.
Apoptosis in CD33+In Fig. 2B, the ∆ caspase-3 value in the MDS group was higher than that in the control group, with a significant p-value. This suggests that the exposure of rhTNFα led to increased apoptosis in both groups, but the impact of increasing apoptosis in CD33 + MDS cell group was more significant.
Raza et al. [11] found that in MDS, exposure of TNFα would result in increased apoptosis in both CD33 + and CD34 + cells. In MDS, despite the excess apoptosis in the bone marrow, granulocyte in the peripheral circulation tend to be more resistant to apoptosis. This occurs because sensitive granulocytes undergo apoptosis earlier in the bone marrow, while apoptosis-resistant granulocytes continue to enter the circulation. Apoptosis in the myeloid lineage is also affected by myeloid-derived suppressor cells (MDSCs). In MDS, there is an increase in circulating MDSCs. MDSCs induce apoptosis in hematopoietic progenitor cells and trigger the secretion of suppressive cytokines, IL10 and TGFβ, which suppress the development of immature myeloid cells [25].
Apoptosis in CD41+In Fig. 2C, caspase-3 activity in CD41 + in the MDS group exposed to rhTNFα was significantly higher compared to the group without exposure. This indicated that TNFα exposure to MDS megakaryocyte-platelet lineage cells leads to increased apoptosis. Cevic et al. [26], also observed that TNFα increased caspase-3 activity in platelets.
Apoptosis in megakaryocyte-platelets lineage of MDS can occur through the extrinsic pathway involving TNFα, but it predominantly takes place via the intrinsic pathway. This is supported by the finding of higher levels of Bax and Bak proteins [27], as well as the possibility of mitochondrial activation or the release of cytochrome C from the cytosol [28].
Following the maximum release of platelets (1000–3000 platelets), senescent megakaryocytes, characterized by a thin nucleus and cytoplasm, immediately undergo apoptosis and are phagocytosed by macrophages [29]. The apoptosis in MDS megakaryocytes occurs due to ineffective platelet production caused by the premature apoptosis of mature megakaryocytes in the intramedullary. Cytogenetic abnormalities in MDS clones primarily led to defects in megakaryocytes, manifesting in morphological abnormalities, and disturbances in nucleus differentiation and maturation. Moreover, these abnormalities are influenced by the MDS bone marrow microenvironment.
To address the hypothesis regarding whether apoptosis in MDS progenitor cell occurs during the early stage or at a differentiation stage, a post hoc analysis was performed to examine the apoptotic activity of caspase-3 in CD34 + cells, representing early progenitors, and in CD33 + and CD41 + cells, representing differentiated progenitors. This study demonstrates that exposure to rhTNFα significantly increases caspase-3 activity, especially in hematopoietic progenitor cells that differentiate into the myeloid lineage CD33 + and the megakaryocyte-thrombocyte lineage CD41+, as opposed to early progenitor lineage CD34+, Fig. 4.
Fig. 4
Scheme of Apoptosis Mechanism on MDS Hematopoesis Progenitor Cell by. TNFα. Early hematopoesis progenitor cell express CD34 + surface marker. Then it will differentiate into specific myeloid progenitor (CD33+), megakaryocyte-thrombocyte progenitor (CD41+). This image illustrates that TNFα induces apoptosis in MDS hematopoesis progenitor cell, especially to cell that have differentiated into myeloid CD33 + and megakaryocyte-thrombocyte CD41+
These findings shed light on why cytopenia in MDS is more pronounced than the increase in blast cells. Similar results were reported by Xia et al. [30], who examined apoptotic activity in MDS CD34+ (progenitor cells) and MDS CD34- (mature cells), and found higher apoptotic activity in mature cells. However, their study did not distinguish the specific stage of differentiation among mature cells.
These results contrast with the previous hypothesis, which suggested that apoptosis in MDS mainly occurs in early progenitor cells CD34+.
The reason MDS develops slowly but progressively lies in the fact that CD34 + MDS cells are long-term repopulating stem cells (LT-HSC) with a high self-renewal capacity but a slow proliferation rate. The MDS cell clone could persist for up to 17 years without clinically significant changes in bone marrow or peripheral blood. However, after prolonged perios, the bone marrow exhibits an 8.15% presence of dysplastic progenitor cells. These abnormal progenitor cells would significantly reduce the number of mature cells in the peripheral blood, leading to cytopenia [31].
This explanation helps clarify the typical phenomenon in MDS, where there is hypercellularity in the bone marrow but cytopenia in the peripheral blood. It suggests that apoptosis occurs mainly in differentiated progenitor cells rather than in early progenitor cells. Unlike other malignant cells that can further develop, MDS clonal cells fail to mature, leading to an accumulation of cells with increased apoptotic abilities.
Comments (0)