Huge evidence indicates that high levels of pro-inflammatory cytokines encourage neuronal malfunction and death, which is why neuroinflammation has been linked to the emergence of neurodegenerative diseases. Therefore, it is crucial to research potential substances that reduce the inflammatory response in the central nervous system. The current study is the first to evaluate the prophylactic anti-neuroinflammatory effects of meclizine against LPS-induced neuroinflammation in mice. This study is also extended to look out for the possible molecular signaling pathways mediating these possible anti-neuroinflammatory actions.
In the current work, the intraperitoneal injection of LPS (5 mg/kg) caused significant neuroinflammation in mice as was revealed by significant elevation of brain tissue contents of IL-1β, TNF-α, NF-κβ, AKT, ERK and JNK. Moreover, significant inflammatory histopathological changes along with positive GFAP immunohistochemical staining have been noticed in mice brains following LPS administration. Meclizine oral prophylactic administration resulted in significant anti-inflammatory effects.
LPS -an endotoxin isolated from the Gram-negative bacterial cell wall- is capable of inducing neuroinflammation, and neurotoxicity along with cognitive impairment in experimental animals. Numerous signaling pathways have been proposed to elucidate LPS-induced neuroinflammation, however, the exact mechanisms have not yet been well-identified (Fasolo et al. 2021). Neuroinflammation is a cascade of inflammatory reactions that occur in the CNS, accompanied by the massive production of cytokines, chemokines and inflammatory enzymes. The CNS contains two main types of immunocompetent cells, viz., microglia and astrocytes. Microglia maintain homeostasis in the CNS where they sense and remove excessive metabolic waste and cellular debris. Likewise, astrocytes play a crucial function in maintaining BBB and are fundamental for both the growing and adult brain. Therefore, both microglia and astrocytes play an essential role in neuroinflammation (Skrzypczak-Wiercioch and Sałat 2022).
LPS administration resulted in significant activation of the microglia. Microglial activation stimulates the triggering of pro-inflammatory mediators such as NF-kβ, TNF-α, IL-16, IL-1β, NO, and reactive oxygen species (ROS) to stimulate tissue repair (Gargouri et al. 2018). It is well established that the NF-κβ pathway is essential for the CNS, especially in cases of brain inflammation, acute cerebral traumas, and neurodegenerative diseases (Mostafa et al. 2021a). NF-κβ is crucial for the microglia-mediated inflammatory reaction in the CNS (Raasch et al. 2011). NF-kβ signaling initiates inflammatory responses via the activation of numerous pro-inflammatory cytokines in most cells (Mostafa et al. 2022). Cytokines are small polypeptides produced by a variety of cells controlling cell growth, differentiation, inflammation and wound healing (Asaad and Mostafa 2022). NF-kβ signaling causes the activation of TNF-α and interleukins. Dysregulation in NF-kβ signaling justifies the pathogenesis of numerous inflammatory syndromes (Mostafa et al. 2021b).
Similar to our work, many studies suggested the involvement of NF-kβ signaling in LPS-induced neuroinflammation (Batista et al. 2019; Guo et al. 2022; Skrzypczak-Wiercioch and Sałat 2022; Yegla and Foster 2019). Gargouri et al. (2018) reported stimulation of TNF-α, IL-1β along with IL-6 in LPS-activated primary microglial cells (Gargouri et al. 2018). Another study reported TNF-α and IL-1β release from both astrocytes and microglia in response to neuroinflammation following LPS administration in mice (Rojas-Colón et al. 2021).
Numerous inflammatory diseases, malignancies and autoimmune conditions stimulate the AKT signaling pathway. It is well-documented that effective NF-kβ signaling and the expression of NF-kβ-regulated inflammatory cascade depend on the AKT activation (Mostafa and Salama 2023). NF-κβ is the main downstream target for AKT (Mostafa and Abdel-Rahman 2023).
Similar to the current work, reported stimulation of AKT by LPS-induced neuroinflammation in LPS-activated microglia (Zhao et al. 2019). A key regulator of IL-6 and TNF-α is ERK. It is noteworthy that ERK production is excessively triggered in several inflammatory and auto-immune disorders (Mostafa and Salama 2023). JNKs are members of the family of mitogen-activated protein kinases that are activated by a variety of stimuli, such as inflammation, oxidative stress, and brain ischemia-reperfusion injury. JNKs are expressed in all cells and tissues throughout the body. JNKs are thought to play a crucial role in the processes involved in neuronal injury and are usually involved in the development of neuroinflammation, stroke, Alzheimer’s disease and Parkinson’s disease. Therefore, JNK inhibitors have potential neuroprotective effects (Anfinogenova et al. 2020). Similar to the current work, Lim et al. (2018) suggested that LPS-induced neuroinflammatory changes in mice brains may be attributed to NF-κB along with several mitogen-activated protein kinases; viz., ERK and JNK (Lim et al. 2018). Consistent data was also reported where mitogen-activated protein kinases signaling, including ERK, and JNK up-regulation initiate numerous neuroinflammatory mediators (Jung et al. 2016; Qi et al. 2016). Similarly, Guo et al. (2022) reported the involvement of the ERK/JNK/NF-κB signaling pathway in LPS-induced neuroinflammation induced experimentally in mice (Guo et al. 2022).
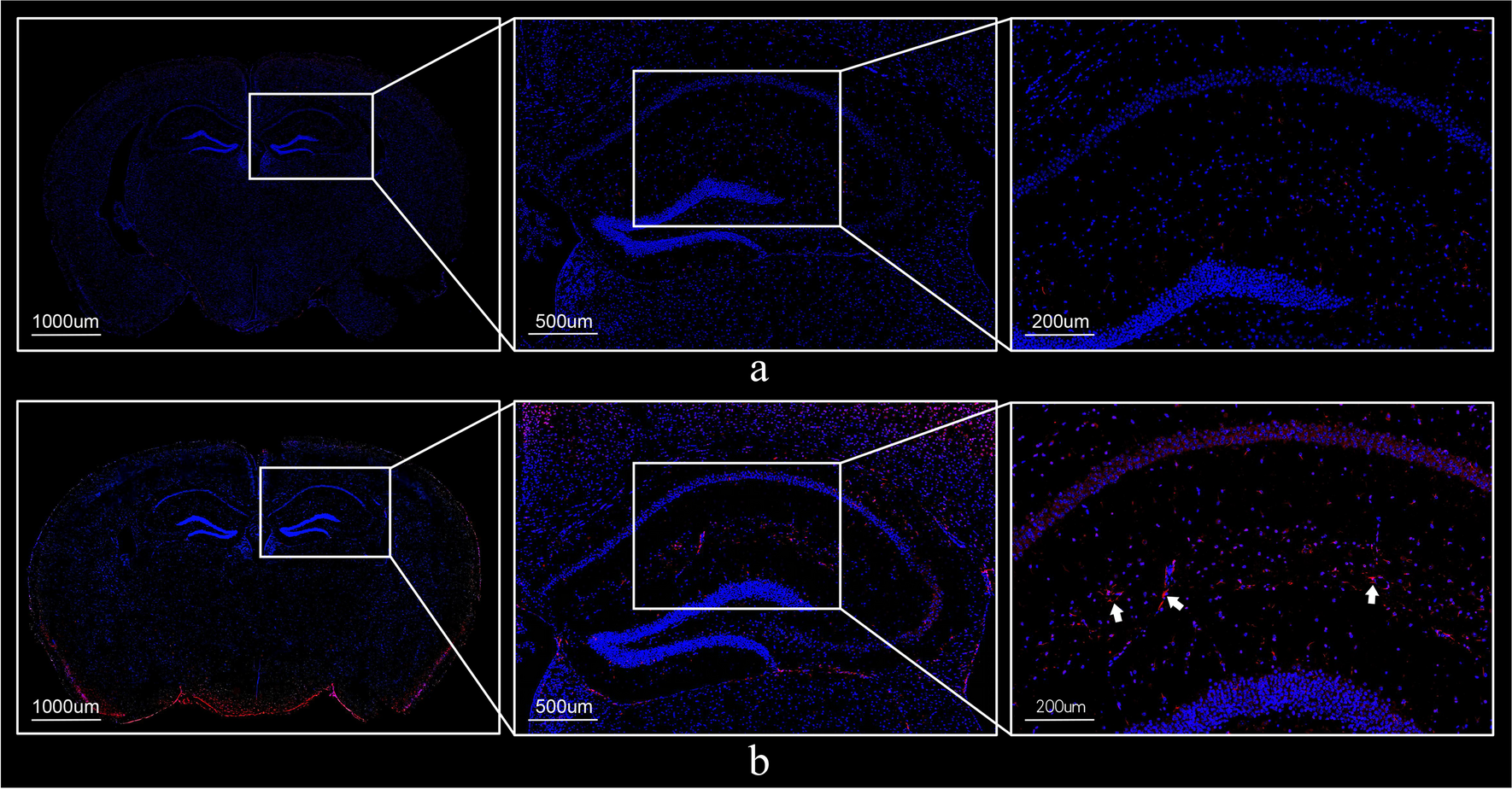
GFAP is an immunohistochemical indicator overexpressed in response to neuronal and astrocyte inflammatory injury. In the context of neuroinflammation, GFAP expression in astrocytes is the key element of the glial scar formation (Mostafa et al. 2021a). The current study reveals a significant elevation in the GFAP immunohistochemical reactivity expressed as reaction area percent in LPS-control mice. In line with this finding, many studies reported that LPS causes abundant cellular proliferation neuroinflammation and GFAP expression in experimental injuries of the CNS in rodents and human cell lines. (Gao et al. 2020; Niranjan et al. 2014).
Meclizine, a histamine antagonist derived from piperazine, is currently used as an “over-the-counter” H1 receptor blocker to reduce motion sickness-related nausea, vomiting, and dizziness. It is a widely prescribed, well-tolerated drug. Meclizine’s neuroprotective properties in a variety of neurological conditions, including ischemic stroke, Parkinson’s disease and Huntington’s disease, have recently come under investigation (Hong et al. 2016).
Meclizine seems to be usually well tolerated overall. Among its most frequent side effects is sleepiness. Numerous investigations concluded that meclizine had little to no effect on perceptual efficiency and detrimental memory (Patel and Ambizas 2011). Studies examining the CNS effects of meclizine 50 mg found that it significantly increased recognition and reaction time (Manning et al. 1992). The US Food and Drug Administration (FDA) declared that there is insufficient evidence to justify restricting the use of meclizine during pregnancy. (Patel and Ambizas 2011). Preclinical safety studies of repeated oral administration of meclizine once or twice a day for 14 days appeared to be safe and well-tolerated with no serious adverse events (Kitoh et al. 2020).
MCLZ use in the current work was linked to significant anti-inflammatory effects as manifested by a statistically significant reduction in brain tissue concentration of brain tissue contents of IL-1β, TNF-α, NF-κβ, AKT, ERK and JNK. Additionally, significant amelioration of the histopathological inflammatory changes and GFAP immunohistopathological staining have been noticed in the brains of mice treated with MCLZ.
MCLZ has been previously reported to cross the blood-brain barrier and to possess cytoprotective and anti-neurodegenerative actions. These effects are independent of its anti-muscarinic or anti-histaminergic actions (Gohil et al. 2011).
Earlier studies reported that MCLZ’s neuroprotective effects comprise suppression of apoptosis along with increased glycolysis without affecting levels of total ATP (Hong et al. 2016).
Singh et al. (2020) stated that MCLZ spares cognitive functions in mice due to its antioxidant and anti-inflammatory actions. MCLZ administration caused a significant reduction in proinflammatory cytokines (IL-1β & TNF-α) in mice brains post streptozotocin administration (Singh et al. 2020). Another study reported that MCLZ decreased mitochondrial oxygen consumption, oxidative stress and inflammation as manifested by reduced IL-6, IL-1β & TNF-α in kidney tissues of mice (Kishi et al. 2015). Guo et al. (2017) reported that MCLZ attenuated ovariectomy-induced bone loss via attenuation of NF-κβ, ERK and p38 (Guo et al. 2017). A recent study also highlighted the possible anti-inflammatory effects of MCLZ where it inhibits the activation of the JNK and NF-κβ and inflammatory cytokines in allergic airway inflammation in mice (Jang et al. 2023).
To the authors’ knowledge, the current work is the first to assess MCLZ’s protective anti-neuroinflammatory effects. The study suggests that MCLZ’s effects are in virtue of the amelioration of the AKT/NF-κβ/ERK/JNK signaling pathway. Based on all the collected results, MCLZ can be of value in protecting against neuroinflammatory disorders. Further investigations are warranted to add MCLZ to the treatment protocol for various neuroinflammatory disorders.

留言 (0)