
記住我
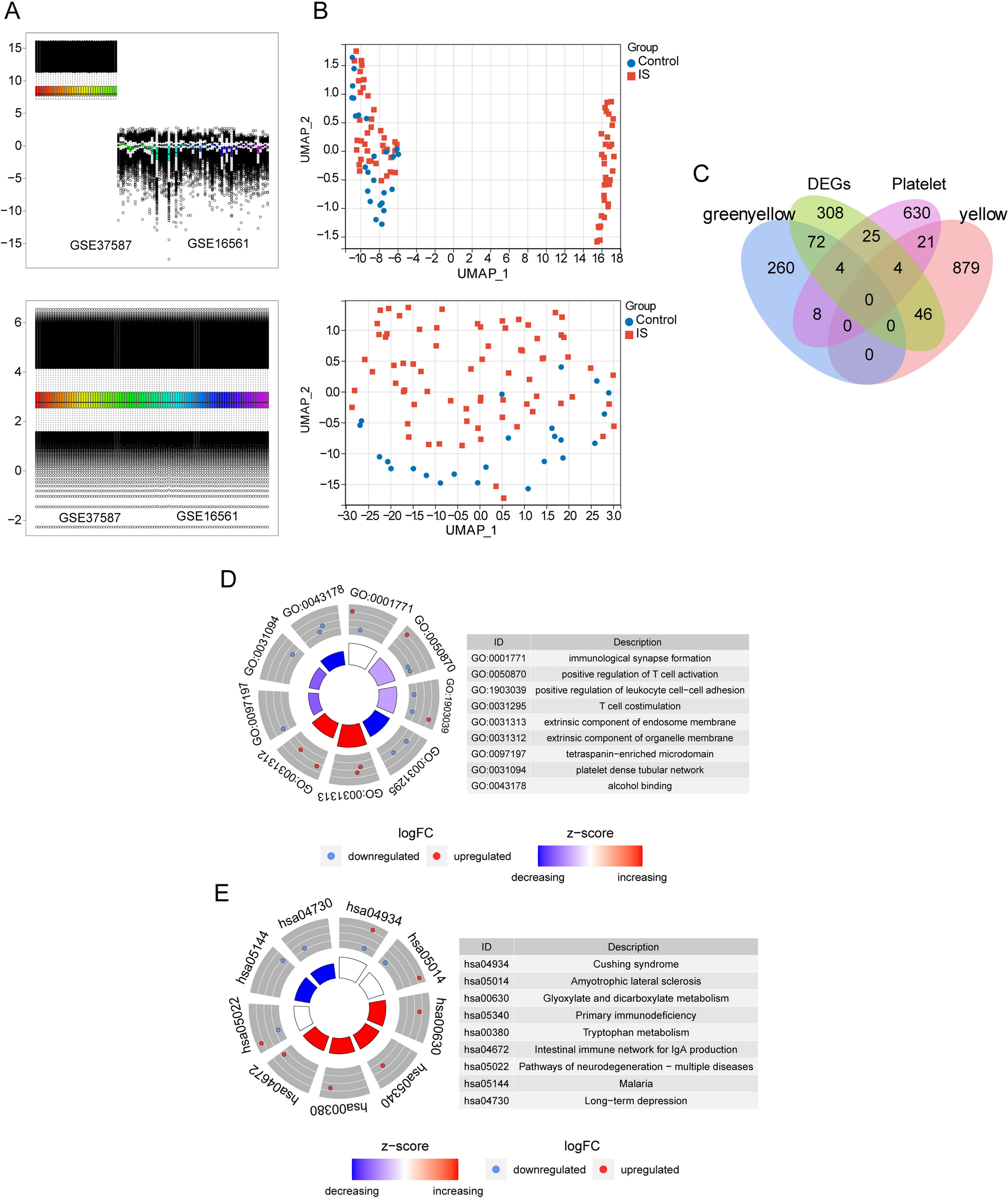
To explore the genetic mechanisms underlying the age-associated diminished regeneration capacity, gene expression profiles in sciatic nerves of aged rats were subjected to dimension reduction using principal component analysis (PCA). The results demonstrated that genes in sciatic nerves of aged rats underwent sham surgery at 1d (A-con-1d), 4d (A-con-4d), and 7d (A-con-4d) gathered together and were clearly separated from aged rats underwent nerve crush injury (A-SNI-1d, A-SNI-4d, A-SNI-7d). The comparison of gene profiles of injured aged rats showed that there seemed to be a gap between gene expressions of aged rats at 1d after nerve injury and gene expressions of aged rats at relative longer time points. Gene expression profiles in aged rats at 4d and 7d after nerve crush injury were far less distinguishing (Fig. 1A). The cooperative display of gene expression profiles of young rats showed that young animals exhibited different expression patterns as aged animals. The long distance between gene expression profiles of uninjured sciatic nerves of aged rats and young rats suggested huge differences between aged and young animals in naïve conditions (Y-con-0d). Still, in young rats, gene expression patterns were similar at longer time points after nerve injury but different from gene expressions at early time points after injury (Y-SNI-1d, Y-SNI-4d, Y-SNI-7d) (Fig. 1A).
Fig. 1
Aged and young animals exhibit unique gene expression patterns post injury. A Principal component analysis (PCA) of young and aged samples. Red dots represent aged animal samples from sham groups (A-con-1d, A-con-4d, A-con-7d) and sciatic nerve injured groups (A-SNI-1d, A-SNI-4d, A-SNI-7d), while green dots represent young animal samples from sham groups (Y-con-0d) and sciatic nerve injured groups (Y-SNI-1d, Y-SNI-4d, Y-SNI-7d). B The number of differentially expressed coding genes (DEGs) between control and injury group in young or aged animals. C Volcano Plot of differentially expressed coding genes in different comparison groups (upper: comparison groups in 1d, middle: comparison groups in 4d, bottom: comparison groups in 7d). Y-axis means log2(fold change). Pink dots represent up-regulated genes and blue dots represent down-regulated genes. D Scatter plots of young or aged preference differentially expressed coding genes after nerve injury. Eight clusters with distinct colors were divided based on the cutoff of fold-change (2, grey dotted lines). upper: comparison groups in 1d, middle: comparison groups in 4d, bottom: comparison groups in 7d. E Importantly enriched GO terms in each cluster in D were shown in different colors. upper: comparison groups in 1d, middle: comparison groups in 4d, bottom: comparison groups in 7d
Given that coding genes are directly related to biological functions, the numbers of differentially expressed coding genes in aged and young rats were summarized. Compared with the corresponding uninjured controls, approximately a total of 5000 genes were differentially expressed in aged rats at 1d and 4d after injury, with approximately 2000 genes up-regulated and 3000 genes down-regulated. A moderately smaller number of genes were differentially expressed, especially down-regulated, in aged rats at 7d after injury. Distinctly different from aged rats, the majority of differentially expressed coding genes in young rats were up-regulated and only a small portion of genes were down-regulated (Fig. 1B). The up and down regulated genes in different comparison groups were displayed in volcano plots in detail (Fig. 1C). Moreover, gene profiles of injured aged rats were contrasted with those of young rats to obtain aged preference differentially expressed coding genes after nerve injury (Fig. 1D). Enrichment of these aged preference differentially expressed coding genes to GO terms indicated that at 1d after nerve injury, genes up-regulated in aged but not young rats (cluster 1 and 2) were mostly enriched in terms related with steroid biosynthetic process (cluster 1) and inflammatory response (cluster 2); genes have similar tendency in aged and young rats (cluster 3 and 7) were mainly associated with inflammatory response, leukocyte activation, positive regulation of response to external stimulus (cluster 3) and glia cell differentiation and extracellular structure organization (cluster 7); genes down-regulated in aged but not young rats (cluster 8 and 9) were mostly enriched in terms related with sensory organ development (cluster 8) and muscle system process (cluster 9) (Fig. 1E).
Inflammatory response and mitotic cell cycle were still enriched in both aged and young up-regulated genes at 4d and 7d after nerve injury and cell junction organization and regulation of membrane potential were enriched in both aged and young down-regulated genes at 4d and 7d after nerve injury. Some other GO terms, such as synaptic signaling and ion transport were emerged as aged down-regulated biological processes at 4d and 7d after nerve injury (Fig. 1E).
Regulation patterns of aged and young rats were further investigated using WGCNA analysis. WGCNA analysis elucidated a total of 20 closely co-expressed gene clusters (modules) (Fig. 2A). Uninjured and injured sciatic nerves of aged and young rats showed diverse WGCNA modules. Lightcyan module was specifically highly involved in the intact sciatic nerves of aged rats instead of injured nerves of aged rats or young rats. Darkgray module, on the contrast, was involved in the injured sciatic nerves of aged rats instead of uninjured nerves of aged rats or young rats. Darkred module was characteristically involved in sciatic nerves of aged rats at 4d and 7d after injury. Besides these specific modules, some modules are commonly enriched in both aged and young rats. Genes in both aged and young rats at 1d and 4d after nerve injury were enriched in darkolivegreen module. Genes in aged rats at 4d and 7d after nerve injury as well as young rats at 1d and 4d after nerve injury were enriched in orangered4 module. Young rats displayed some unique features. For instance, compared with aged rats, genes in both uninjured and injured young rats, especially injured young rats, were enriched in darkmagenta module (Fig. 2B).
Fig. 2
Weighted gene correlation network analysis (WGCNA) analysis reveals differential regulation patterns in aged and young post injury. A WGCNA clustering dendrogram. Co-expressed genes were clustered in the same module and labeled with different colors. B The correlation between modules and groups. C Heatmap showing relative expression of genes in lightcyan, darkgrey, darkred, darkolivergreen, orangedred4, and darkmagenta modules identified by WGCNA of transcriptomes in young and aged animal after nerve injury. Representative Gene ontology (GO) terms of each significantly-regulated module are listed below. D, E Boxplots showing expression patterns (scaled log2TPM) of hub gene (top 10) in darkolivegreen (D) and orangered4 (E) module
Genes in these critical WGCNA modules were demonstrated in a heatmap and functionally explored. Genes belonging to lightcyan module, mainly genes in the uninjured aged sciatic nerves, were associated with ion transport, system development, and lipid metabolic process. Genes belonging to darkgrey module, that is chiefly genes in the injured aged sciatic nerves, were associated with ion transport, antigen processing and presentation, organic substance, and catabolic process. Highly related biological processes in other WGCNA modules were discovered as well, including system development, response to external stimulus, and synapse pruning in darkgrey module, positive regulation of immune system process, cell activation, and cytokine production in darkolivegreen module, cell cycle, DNA metabolic process, and cell division in orangered4 module, and chromosome organization, cell macromolecule metabolic process, and cell cycle process in darkmagenta module (Fig. 2C). Darkolivegreen and orangered4 modules attracted most attention as gene belongs to these two modules were robustly differentially expressed after nerve injury for both aged and young rats and showed temporal specificity. Additional exhibition of top 10 genes in darkolivegreen and orangered4 modules showed that consistent with all genes in these two modules, hub genes were expressed at higher levels after nerve injury in both aged and young rats (Fig. 2D, E).
Identification of Robust Immune Responses in Aged and Young Animals After Peripheral Nerve InjuryA full demonstration of genes in WGCNA darkolivegreen module indicated that as compared with their corresponding uninjured control, in both aged and young rats, the expression levels of genes in darkolivegreen module increased to a log2RPKM + 1 value of approximately 6 at 1d after nerve injury. The expressions of the majority of genes in darkolivegreen module were slightly decreased at 4d and 7d, especially in young rats. Still, the expressions of these genes remained at high levels as compared with the uninjured controls (Fig. 3A). GO analysis showed that genes in darkolivegreen module were primarily associated with immune responses and immune response-regulated activities (Fig. 3B). The substantial involvement of immune responses was identified by KEGG analysis as well, with B cell receptor signaling pathway (Ko04662) being the most significant KEGG pathway (Fig. 3C). Many genes that participate in the antigen-receptor signals of B cells, including CD22 (CD22 Molecule), LYN (LYN proto-oncogene, Src family tyrosine kinase), BTK (Bruton agammaglobulinemia tyrosine kinase), SYK (Spleen Associated Tyrosine Kinase), Bam32 (dual adaptor for phosphotyrosine and 3-phosphoinositides 1), PLC-γ2 (phospholipase C, gamma 2), SHIP (also called INPP5D, Inositol Polyphosphate-5-Phosphatase D), and BCAP (also called ODF2L, Outer Dense Fiber Of Sperm Tails 2 Like), were elevated in both aged and young rats after nerve injury (Fig. 3D).
Fig. 3
Aged and young animals have different immune cell responses post injury. A Fold changes of genes in darkolivegreen module. Red lines represent fold changes of genes in young animals and green lines represent fold changes of genes in aged animals. B, C: The enriched GO terms (B) and Kyoto Enrichment of Genes and Genomes (KEGG) pathways (C) of genes in darkolivegreen module. The most significant KEGG pathway was labeled with red star. D The schematic network of B cell receptor signaling pathway (Ko04662). Up-regulated genes after injury were marked with red box. Signaling pathway diagram was adopted from KEGG database
Given that immune responses are cooperatively regulated by various types of immune cells, we compared immunoinfiltration through the proportion of different immune cells in each group (Fig. 4A). In general, immune cells exhibited dissimilar distributions even in the intact sciatic nerves of aged and young rats. Firstly, there was a significant proportion of CD8 + T cells in aged rats both in shamed and injury groups while the proportion of CD8 + T cells in control group in young rats was very low. The expression of Cd8 was very low in uninjured young rats when compares with aged animals (Fig. 4A and B). This phenomenon was consisted with the report before in DRGs of aged rats (Zhou et al. 2022), indicated the chronic inflammation in aged animal. The proportion of memory resting T cells in the uninjured young sciatic nerve was obviously much larger while the proportion of B cells was noticeably much smaller as compared with the uninjured sciatic nerves of aged rats (Fig. 4A and B). To conform these results, we explored the expression of IGM, a marker of B cells, through immunostaining in the sciatic nerve of both old and young rats before and after injury. We found that in the absence of injury, the sciatic nerve of old rats expressed more IGM than that of young rats, but after injury, the IGM of both old and young rats continued to increase (Fig. 4D, E), which just consistent with immune infiltration analysis.
Fig. 4
The immunoinfiltration of aged and young rats after sciatic nerve injury. A The proportion of different immune cells in different groups. B The expressions of markers of different types of immune cells in different groups. C The immunostaining of immunoglobulin heavy chain (IGM) in sciatic nerve of aged rat before or after injury. Red represents IGM and blue represents DAPI (cell nucleus), the scale bar = 100 μm. D The immunostaining of immunoglobulin heavy chain (IGM) in sciatic nerve of young rat before or after injury. Red represents IGM and blue represents DAPI (cell nucleus), the scale bar = 100 μm. E: IGM signal intensity statistics, Y axis represents the average gray value of IGM signal, X axis represents different groups, n = 3
Following sciatic nerve injury, larger amounts of dendritic cells and macrophages were observed in both aged and young rats, with increased macrophages occurred at early time points after nerve injury and increased activated dendritic cells occurred at later time points after nerve injury (Fig. 4A). Aged and young rats also displayed specific immune responses to nerve injury. In aged rats, increased proportion of neutrophils and elevated expression of its marker Cxcr2 (C-X-C Motif Chemokine Receptor 2) as well as ip-ncreased proportion of regulatory T cells (Tregs) and elevated expression of its maker Ltb (Lymphotoxin Beta) were observed at 1d after nerve injury. Higher level of Acp5 (Acid Phosphatase 5, Tartrate Resistant) and larger amount of resting dendritic cells was identified in aged rats at 4d and 7d after nerve injury. Compared with aged rats, in young rats, increased proportion of Monocytes and elevated expression of its marker Mnda (Myeloid Cell Nuclear Differentiation Antigen) at 1d after nerve injury (Fig. 4A and B). These findings demonstrated that aged and young rats underwent robust but unique immune responses.
Identification of Activated Cell Cycle in Young Animals After Peripheral Nerve InjuryThe dynamic expression patterns of genes in orangered4 module, another WGCNA module significantly involved nerve injury-mediated expression changes, were displayed in details as well. For both aged and young rats, genes in orangered4 module increased to a similar level at 4d after nerve injury. However, compared with aged rats, in young rats the immediate injury response of genes in orangered4 module was more robust and genes increased to a higher level at 1d after nerve injury (Fig. 5A). These differentially expressed genes were closely associated with cell cycle, chromosome segregation, nuclear division, DNA metabolism, DNA replication, and cell division (Fig. 5B).
Fig. 5
Cell cycle is more active in young animal post injury. A Fold change of genes in orangered4 module. Red lines represent fold change of genes in young animals and green lines represent fold change of genes in aged animals. B The enriched GO terms of genes in orangered4 module. C The ranking of transcription factors based on the connectivity within the orangered4 module. The size of plot represents gene expression at 1d after injury. D The expressions of transcription factors within orangered4 module, the grey lines indicated interaction between transcription factors and targets. E The interactions between transcription factors and targets within orangered4 module. The transcription factors were labeled with cyan diamond and target were labeled with pink circle. The grey lines indicated interaction between genes which were predicted from IPA software. F The expressions of the targets of these transcription factors within orangered4 module
To explore the switch of these cell cycle related genes, the connectivity and expressions of transcription factors were investigated. There are 9 transcription factors in orangered4 module, with Foxm1 (Forkhead Box M1) have highest connectivity and Msc (Musculin) have lowest connectivity (Fig. 5C). In young rats, most transcription factors and target genes were rapidly up-regulated in 1d after nerve injury while in aged rats, transcription factors and target genes showed increased trends at a later time point, that is 4d after nerve injury. Among these differentially expressed transcription factor, Hmga1 (High Mobility Group AT-Hook 1), Foxm1, Wdhd1 (WD Repeat And HMG-Box DNA Binding Protein 1), E2f7 (E2F Transcription Factor 7) and Nsd2 (Nuclear Receptor Binding SET Domain Protein 2) were first increased and shown higher expression in young rats after nerve injury while LOC102552640, Mybl2 (MYB Proto-Oncogene Like 2) and Cenpo (Centromere Protein O) have higher expression at 4d after injury in aged rats (Fig. 5D).
The targets of these transcription factors and the relationship between them were further explored. Foxm1 appears as the hub of the network, and Ccnb1 (Cyclin B1) also shown higher connectivity among the targets (Fig. 5E). The expression of these targets shown similar to the transcription factors that most of them were rapidly up-regulated in 1d in young but not in aged rats. Among the targets, Pttg1 (PTTG1 Regulator of Sister Chromatid Separation, Securin) and Ezh2 (Enhancer of Zeste 2 Polycomb Repressive Complex 2 Subunit) have little up-regulated after injury in aged rats, but shown obvious higher expression after injury in young rats. Although the expression of Bub1b (BUB1 Mitotic Checkpoint Serine/Threonine Kinase B), Ccne1(Cyclin E1), Cdc25c (Cell Division Cycle 25C) and Gins4 (GINS Complex Subunit 4) were both increased after injury in young and aged rats, they appear higher expression in young animal. In addition, some genes like Cks2 (CDC28 Protein Kinase Regulatory Subunit 2) and Plk1 (Polo Like Kinase 1) shown little change at 1d but rose rapidly at 4d after injury in aged rats compared with young rats (Fig. 4D).
留言 (0)