
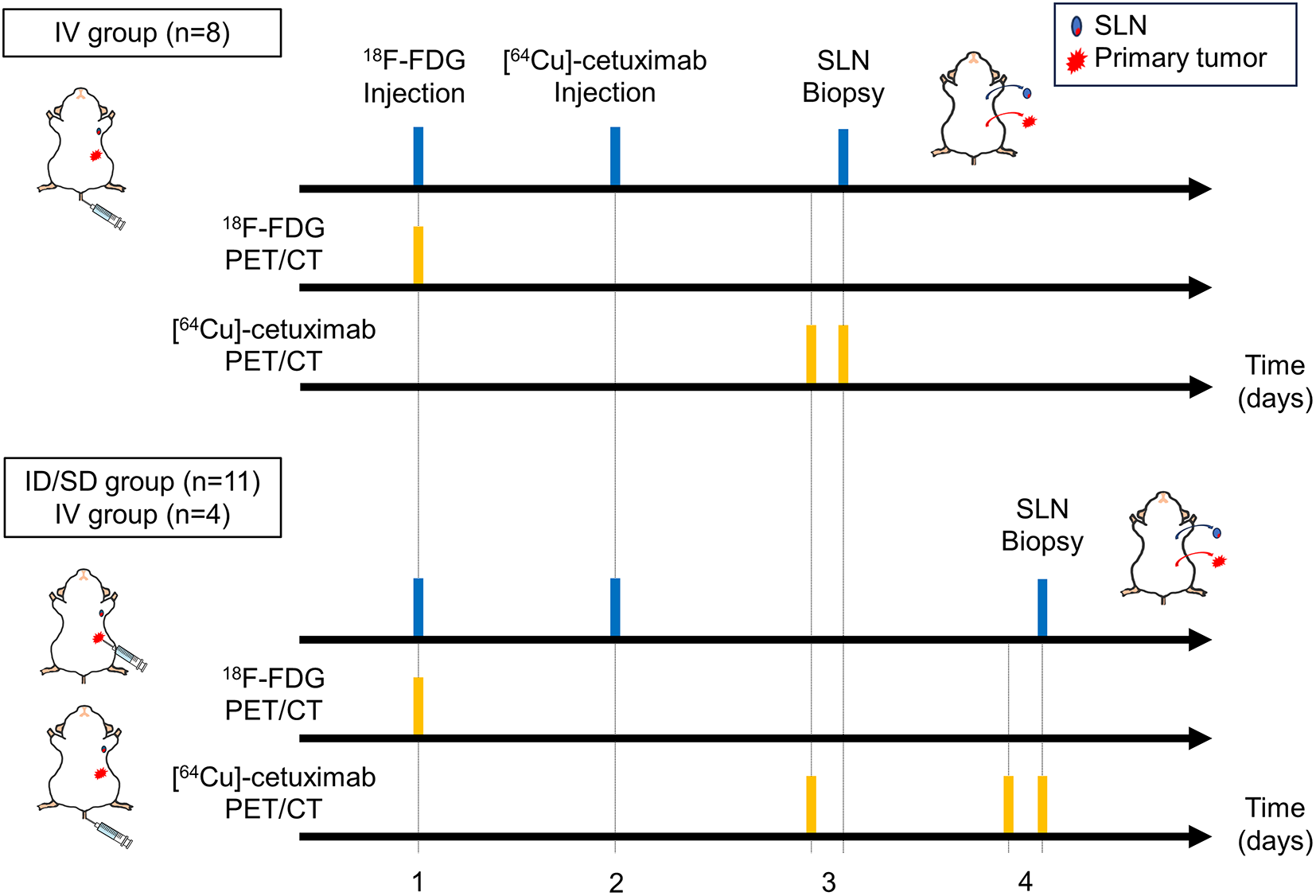
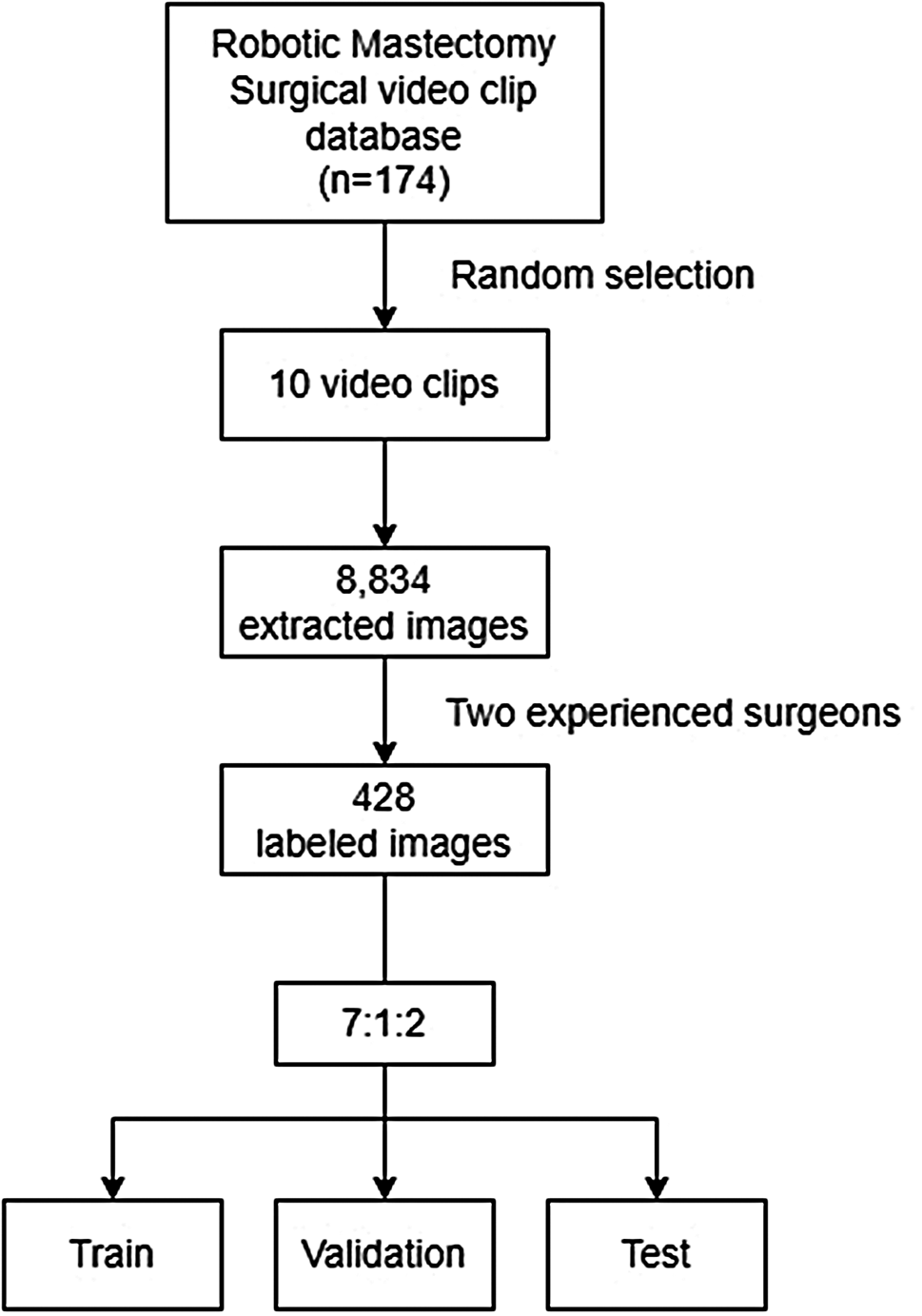
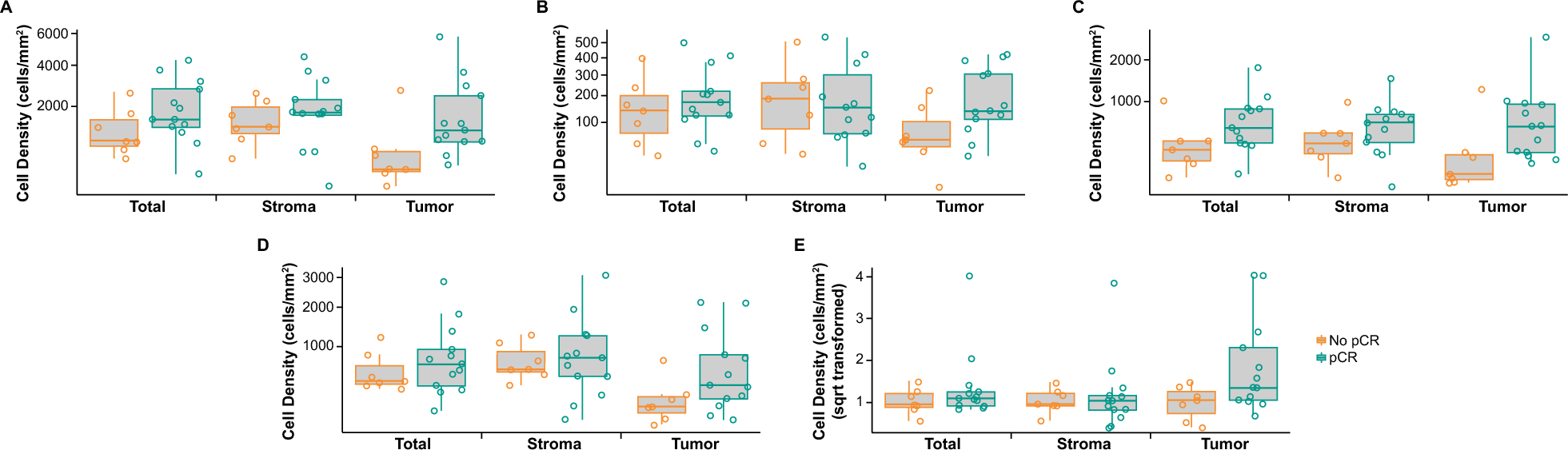
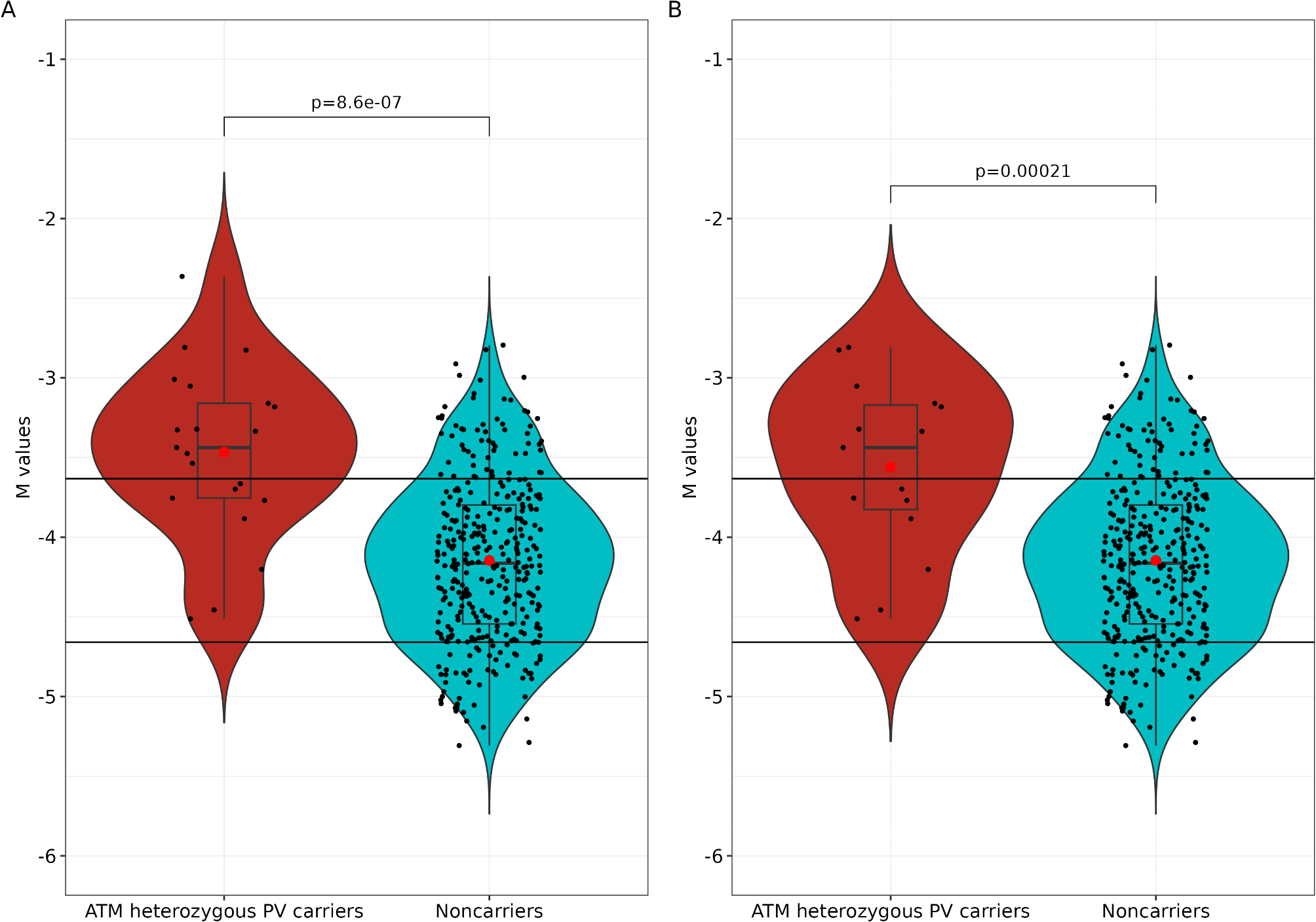
Remember me






Expression of both the transcription factor SNAIL1 and the fibronectin isoform including the EDA domain in myofibroblasts has been described [20, 39]. To determine whether SNAIL1 is involved in the controlling EDA exon inclusion, we first used RT-PCR to visualize the relative amount of skipping/inclusion isoforms in RNA from control or Snai1-deficient fibroblasts, including CAFs, MEFs and human BJ fibroblasts. Oligonucleotide primers annealing to the flanking EDA exons were used at not saturating cycles. We observed a qualitative switch toward EDA-skipped RNA, confirmed by densitometric quantification of the bands, in Snai1 KO relative to control CAFs (Fig. 1A). As described, we detected that the cytokine TGFβ promoted EDA+ fibronectin RNA expression and that the increase was SNAIL1 dependent (Fig. 1B; Additional file 1: Fig. S1). By analyzing EDA+ fibronectin expression at the protein level using a specific monoclonal antibody, we found that it was only faintly detected in Snai1 KO fibroblasts as compared to control cells (Fig. 1C). In human BJ fibroblasts, depletion of SNAIL1 using a specific siRNA downregulated EDA+ fibronectin levels in both untreated and TGFβ-treated cells (Fig. 1D) indicates that the Snai1-depletion effect was not limited to murine MEFs and CAFs or to the constitutive removal of the factor.
To elucidate whether SNAIL1 extensively controls alternative splicing, we evaluated splicing events by deep sequencing of mRNA from control and Snai1 KO MEFs treated for 3 h with TGFβ. The SUPPA2 and SANJUAN pipelines revealed 674 and 299 significantly different splicing events, respectively, that affected more than 500 genes (Additional file 1: Fig. S2A, Tables S1 and S2). Among these, we found several genes involved in actin cytoskeleton regulation, such as Myl6, AnIn, Macf1, Tpm1, Tpm2, PP1R12A, FlnC and FlnB, as well as ECM genes, such as Col5α1 and FN1; we confirmed some of these genes by RT-PCR (Additional file 1: Fig. S2B). For fibronectin, the number of RNA-seq reads for the events including and excluding EDA allowed us to quantify the relative amount of EDA+ fibronectin in each sample, which was over 75% in control MEFs but was reduced by approximately half in Snai1 KO cells (Fig. 1E).
SNAIL1 protein levels correlate with EDA+ fibronectin RNA expression in advanced cancersIn primary breast tumors, the presence of myofibroblatic CAFs has been associated with tumor progression [5, 6, 10, 41], and the expression of fibroblastic SNAIL1, with a poor prognosis [10]. Here, we analyzed whether the simultaneous expression of SNAIL1 and EDA+ fibronectin correlates with advanced breast tumors. We collected available SNAIL1 protein data (cBioportal; TCGA, Firehose Legacy) and calculated the percentage of EDA + fibronectin RNA (using the web tool TSVdb for the TCGA splicing variants) from 809 invasive breast carcinomas. We categorized tumors according to their stage (I-II or III-IV) and calculated the relative levels for each molecule using a cutoff value (Material and Methods). Overall, 55% of advanced tumors (stages III and IV) expressing high levels of SNAIL1 also expressed high levels of EDA+ FN1. This percentage decreased to 33–37% in the remaining categories (Fig. 2A). In protein extracts from patient-derived xenografts (PDX) corresponding to HER2 + or triple-negative breast neoplasms, the percentage of tumors with high EDA+ fibronectin expression was increased in the PDXs expressing high levels of SNAIL1 (Fig. 2B). Remarkably, we found similar associations when we analyzed available data from other solid tumors in which the stromal component has been shown to be relevant for tumor progression, such as skin cutaneous melanoma [42], lung adenocarcinoma [43, 44] and kidney renal clear cell carcinoma [45] (Fig. 2B–D). Even though available data do not discriminate between tumor and stroma expression, our analysis shows that the expression of two myofibroblast-associated proteins mostly present in the stroma of colon and breast tumors was associated with advanced tumors.
Fig. 2
Elevated percentages of EDA+ fibronectin correlate with high SNAIL1 levels in advanced human tumors. A Breast adenocarcinoma; C kidney renal clear cell carcinoma; D skin cutaneous melanoma; and E lung adenocarcinoma. The percentage of EDA+ fibronectin RNA and the SNAIL1 protein level in each specimen were obtained from the TSVdb and cBioPortal databases, respectively. Tumors were classified according to the levels (low or high) of SNAIL1 and EDA+ fibronectin (see Materials and Methods), and the percentages of high or low EDA+ fibronectin tumors were plotted for each SNAIL1-expressing category. Tumors at the initial (I and II) or advanced (III and IV) stages were analyzed separately (see Materials and Methods). Numbers within the bars indicate the percentage of tumors with high levels of EDA+ fibronectin. When available, data for normal tissue (NT) are also shown. n, number of tumors per group. B Relative protein levels of EDA+ fibronectin and SNAIL1 in PDXs. EDA+ fibronectin and SNAIL1 levels in 29 PDX protein extracts from HER2+ or triple-negative breast neoplasms were densitometrically estimated from Western blots (see Materials and Methods). PDXs were classified according to their levels (low or high) of SNAIL1 and EDA+ fibronectin (see Materials and Methods) and analyzed as in A. n, number of PDXs
SRSF1 binding to exon 33 RNA of fibronectin is SNAIL1-dependentTo further analyze how SNAIL1 controls the EDA inclusion, we focused on SRSF1, a splicing factor involved in controlling this event [34, 35]. Although SRSF1-mediated splicing can be regulated by factor availability [34], we did not detect decreased SRSF1 protein levels or changes in its subcellular localization in TGFβ-activated Snai1 KO relative to control MEFs (Fig. 3A,B). In contrast, we observed that SRSF1 and SNAIL1 colocalized in nuclear granules (Fig. 3B) and co-immunoprecipitated (Fig. 3C), suggesting a more direct molecular connection.
Fig. 3
SRSF1 interactions with the fibronectin EDA exon is regulated by SNAIL1. A SRSF1 protein amount in Snai1 KO MEFs. Control and Snai1 KO MEFs were lysed in SDS buffer after the indicated times of TGFβ1 treatment (5 ng/mL), and protein levels were analyzed by Western blotting. B SNAIL1 and SRSF1 colocalize in the nucleus of MEFs. Control and Snai1 KO MEFs were grown on glass coverslips, treated with TGFβ1 for 24 h and fixed with 4% PFA. The cellular distributions of SNAIL1 and SRSF1 were analyzed by immunofluorescence with specific antibodies. Images were obtained by confocal microscopy. Phalloidin (pink) and DAPI (blue) staining corresponding to depicted cells are shown into a box. Merge images in control MEFs were produced with ImageJ, and colocalization is shown in yellow. C The SNAIL1 and SRSF1 interaction is RNA dependent. Extracts of MEFs treated with TGFβ1 for 3 h were obtained in RIPA buffer, and half of the sample was treated with 400 µg/mL RNase A. RT-qPCR for total fibronectin confirmed the complete elimination of RNA in the samples. Immunoprecipitation was performed using an antibody specific for SRSF1 and agarose beads. Immunoprecipitated proteins were analyzed by Western blotting. D SNAIL1 does not bind to the fibronectin exon 33 RNA. RNA immunoprecipitation (RIP) was performed using an antibody specific for SNAIL1 or an unspecific IgG in samples of MEFs transfected to overexpress Snai1-HA (Additional file 1: Fig. S3) and treated with 5 ng/mL TGFβ1 for 3 h. RNA enrichment in the immunoprecipitates was analyzed by RT-qPCR using primers for exon 33. Bars show binding enrichment as compared to immunoprecipitation using IgG. E SRSF1 binds to the fibronectin exon 33 RNA in a SNAIL1-dependent manner. RIP was performed using an antibody specific for SRSF1 in samples of MEFs transfected to overexpress Snai1-HA (Additional file 1: Fig. S3), or of MEFs KO for Snai1 treated with 5 ng/mL TGFβ1 for 3 h. RNA enrichment in the immunoprecipitates was analyzed by RT-qPCR using primers for exon 33 or HPRT (as a control). Bars show binding enrichment compared to immunoprecipitation using unspecific control IgG. At least three replicates were performed for each immunoprecipitation. F and G SRSF1 and SNAIL1 bind to the EDA coding region in a TGFβ-dependent manner. ChIP was performed with an antibody specific for SRSF1 (F) or SNAIL1 (G) in samples of MEFs transfected to overexpress SNAIL-HA that were treated or not with TGFβ1 for 3 h. Precipitated DNA was analyzed by qPCR using primers targeting Fn1 promoter (+ 116/ + 265), Fn1 exon 7 and Fn1 exon 33 (EDA). Bars show binding enrichment as compared to immunoprecipitation using unspecific IgG. At least three replicates were performed for each immunoprecipitation
As SRSF1 was described to bind EDA RNA [46], we tested whether SNAIL1 influences SRSF1 binding to exon 33 RNA using RNA immunoprecipitation (RIP) assays in Snai1 KO MEFs. SRSF1 or SNAIL1 was immunoprecipitated with specific antibodies, and the co-precipitating RNA was analyzed. Fibronectin RNA did not significantly co-precipitate with anti-SNAIL1, even in MEFs overexpressing ectopic SNAIL1-HA treated with TGFβ (Fig. 3D). RIP using anti-SRSF1 in these cells confirmed that SRSF1 interacts with EDA RNA, as previously reported (Fig. 3E). Remarkably, the binding of SRSF1 to the EDA RNA region (but not to a control irrelevant region, HPRT) was undetected in Snai1 KO MEFs (Fig. 3E). This result indicates that SNAIL1 is required for the specific binding of SRSF1 to the exon 33 RNA.
Splicing factors of the SRSF family are involved in coupling RNA Pol II transcription to pre-mRNA splicing [47] and precipitate genomic DNA in the presence of crosslinking agents [48]. Therefore, we used ChIP to test whether SRSF1 interacts with the genomic DNA at exon 33. We detected that TGFβ promoted precipitation of this region, but not of the fibronectin proximal promoter or exon 7 regions, with SRSF1 (Fig. 3F), indicating that the splicing factor is likely involved in a co-transcriptional complex. As SNAIL1 interacts with SRSF1, we tested whether SNAIL1 also interacts with the exon 33 genomic region. In contrast to the lack of its binding to RNA, we found that SNAIL1 interacted with the exon 33 in a TGFβ-dependent manner, as well as with the proximal FN1 promoter (as previously described; [49]) (Fig. 3G). Lack of binding to the fibronectin exon 7 region supported the specificity of these ChIP interactions.
Altogether, our data suggest that TGFβ induces localization of SNAIL1 at the alternative splicing region, which is required for the formation of a co-transcriptional complex that includes SRSF1 bound to the EDA RNA. In this case, we would expect that the DNA and RNA binding machineries (including SNAIL1 and SRSF1, respectively) connected by nascent RNA, would be separated by an RNase treatment. Indeed, co-immunoprecipitation of SNAIL1 and SRSF1 was disrupted when input lysates were treated with RNase (Fig. 3C).
As the reduction of the EDA-containing isoform in untreated Snai1 KO MEFs (Fig. 1A–C) could be independent of SRSF1, we tested the levels of other splicing factors involved in the regulation of EDA splicing, such as SFSR3, SRSF5 [20, 50] and QKI [51]. While the levels of the splicing silencer QKI increased in the KO MEFs (Additional file 1: Fig. S4A), no clear changes were observed for SRSF1 (Fig. 3A), SRSF3 or SRSF5 (Additional file 1: Fig. S4B). In contrast, in epithelial cells that did not express endogenous SNAIL1 or EDA+ fibronectin, exogenous expression of SNAIL1-HA promoted the increase in EDA+ fibronectin, as well as of SRSF5 and (to a lesser extent) SRSF3 (Additional file 1: Fig. S5A and B).
Fibronectin EDA sustains structural ECM properties regulated by SNAIL1Our previous research has shown that the extracellular anisotropy generated by CAF lines correlates with their SNAIL1 levels and that TGFβ-treated fibroblasts produce anisotropic matrices in a SNAIL1-dependent manner [10]. Analogously, we found here that our CAF line but not the Snai1 KO one generates anisotropic three-dimensional extracellular matrices (3D ECMs). In detail, IF analysis showed aligned fibronectin fibers that decrease in Snai1 KO ECMs (Fig. 4A) and angle measurement of DAPI-stained nuclei indicated that the high proportion of oriented nuclei were decreased by half (Fig. 4B).
Fig. 4
Fibronectin EDA determines topological and mechanical properties of myofibroblastic matrices. A CAF-derived 3D ECMs. Indicated CAFs were seeded on coverslips and allowed to produce ECM for 6 days. Cell cultures were then fixed and analyzed by immunofluorescence (IF) with an anti-fibronectin (red), anti-fibronectin EDA (green) and DAPI (white). B Quantification of the oriented CAF nuclei within 3D ECMs. Orientation angles of the DAPI-stained nuclei were calculated using the ImageJ analysis particles tool. Percentage of nuclei orientated toward the most frequent angle (up to 21° deviation) is shown. C Relative RNA amount of EDA+ fibronectin isoforms in genetically modified MEF lines. RNA from control, EDA– and EDA+ MEFs treated with 5 ng/mL of TGFβ1 for 3 h was retrotranscribed and amplified using primers flanking exon 33 of Fn1 (as described in Fig. 1) and visualized by DNA-electrophoresis. D Relative protein amount of EDA+ fibronectin isoforms in genetically modified MEF lines. Indicated MEFs treated as in C were lysed in SDS buffer, and the levels of the indicated proteins were analyzed by Western blot. E Fibronectin fibers in 3D ECMs. Indicated MEFs seeded on coverslips were allowed to produce extracellular matrix for 6 days in the presence or absence of 5 ng/mL TGFβ1. Cell cultures were then fixed and analyzed by IF with an anti-fibronectin (green) and DAPI. Confocal and STED microscopy were used to obtain images. F Quantification of fibronectin fiber alignment in 3D ECMs. The fiber angles were calculated using the ImageJ plugin OrientationJ. The percentage of fibers aligned toward the same direction (up to 21° deviation from the mode) is shown. G Quantification of fibronectin fiber alignment index through TWOMBLI. Fibronectin fiber images obtained as in E were analyzed using the ImageJ macro TWOMBLI. All obtained data are plotted, showing all individual measurements, mean and SEM. H Quantification of fibronectin fiber parameters through TWOMBLI. The indicated parameters were analyzed from images used in E. Arbitrary units provided by the plugin are expressed relative to wild-type MEFs. I Visualization of collagen deposition from in vivo–like extracellular matrices. 3D ECMs were produced as in C fixed with 4% PFA and stained with Masson's trichrome. J, Quantification of the stiffness of in vivo–like extracellular matrices. 3D ECMs generated as in E were decellularized, and the elastic modulus was calculated from atomic force-curve measurements
Moreover, we observed a reduction of the presence of EDA+ linear fibronectin fibers in ECMs from KO CAFs (Fig. 4A). Therefore, we analyzed the contribution of the EDA+ fibronectin on the extracellular topology using TGFβ-activated MEFs derived from two genetically engineered mouse strains that exclusively expressed either EDA-including (EDA+) or -excluding (EDA–) fibronectin isoforms [19]. RNA analysis confirmed the expression of the corresponding isoforms in these cell lines (Fig. 4C). In control MEFs under standard cell culture conditions, the expression of EDA+ fibronectin was predominant (Figs. 1A,B,E and 4C). Proteins corresponding to EDA+ fibronectin were detected only in the control and EDA+ MEFs (Fig. 4D). 3D ECMs from these MEF lines were produced and analyzed by immunofluorescence with an anti-fibronectin antibody recognizing both EDA+ and EDA– isoforms. The absence of EDA+ fibronectin in EDA– 3D ECMs was confirmed by immunofluorescence (Additional file 1: Fig. S6). In the absence of TGFβ, all lines produced randomly oriented fibronectin fibers (Fig. 4E). Fiber orientation was estimated to be over 33% with the OrientationJ plugin (Fig. 4F), as reported for unaligned fibers deposited by control fibroblasts [9, 10]. In the presence of TGFβ, the increase of oriented fibronectin fibers was significantly lower in matrices produced by EDA– MEFs (Fig. 4E,F), indicating that the presence of the EDA within the matrix favors TGFβ-induced aligned polymerization of fibronectin fibers. High-resolution images obtained by STED (stimulated emission depletion) microscopy illustrated the differences in the fiber nets between the three matrices (Fig. 4E).
A higher fiber alignment in EDA+ relative to EDA– matrices (Fig. 4G), as well as a lower curvature and branch points (Fig. 4H), was confirmed with the TWOMBLI plugin for FIJI [52]. Additionally, differences between the control and EDA+ matrices affecting alignment, curvature, branch points, lacunarity and endpoints were also detected (Fig. 4H). As extracellular fibronectin acts as a template to guide the polymerization of other extracellular fibers, we examined whether collagen deposition was dependent on the fibronectin isoforms. At a glance, the collagen pattern was clearly different in the EDA– matrices (Fig. 4I). As collagen organization and crosslinking determine the rigidity of the ECM, we used atomic force microscopy (AFM) to measure the micromechanical properties of decellularized matrices generated by TGFβ-treated MEFs; this revealed a significantly lower elastic modulus in matrices derived from EDA– fibroblasts (Fig. 4J). Therefore, we conclude that the presence of the EDA in fibronectin is necessary for activated fibroblasts to generate matrices with high alignment and rigidity.
Deposition of fibronectin fibers is sensitive to the action of metalloproteinases in the absence of EDAThe ECM in tumors is deposited in the presence of tumor cells, and matrix-remodeling enzymes, such as metalloproteinases, are activated by signaling resulting from the interactions between tumor cells and fibroblasts [53]. Thus, we evaluated the fibronectin fiber organization in co-cultures of tumor cells and fibroblasts expressing different fibronectin isoforms. In the presence of pre-seeded, Ras-transformed EpH4 breast cells (EpRas), MEFs deposited fibronectin fiber around tumor cell patches visualized as a net of fibers in a fibronectin staining (Fig. 5A). Lacunas between fibers measured by TWOMBLI were larger in co-cultures with MEFs expressing EDA– fibronectin than with the other MEF lines (Fig. 5B). Similar differences were observed in co-cultures with other tumor cells, such as MCF7 (breast) or HepG2 (liver) (Additional file 1: Fig. S7A and S7B). TWOMBLI analyses also detected differences in fiber branchpoints between co-cultures (Additional file 1: Fig. S7C and S7D). Notably, addition of the metalloproteinase inhibitor GM6001 reverted the lacunarity and branchpoints parameters in co-cultures with EDA– fibronectin (Fig. 5A,B, Additional file 1: Fig. S7C).
Fig. 5
Matrices deposited from both EDA– fibronectin and Snai1 KO MEFs are sensitive to metalloproteinases. A Fibronectin fiber organization in EpRas cells co-cultured with fibroblasts. EpRAs cells and the indicated MEFs were co-cultured on glass coverslips in the presence or absence of 25 µM GM6001 for 3 days. Co-cultures were analyzed by IF with anti-fibronectin (green) and DAPI (blue). Microscopy images are shown. B Fibronectin fiber lacunarity in EpRas co-cultured with fibroblasts is EDA and metalloproteinase dependent. Lacunary in fibronectin images obtained as in A was quantified using the TWOMBLI plugin of ImageJ software. The fold-increase with respect to values in untreated control MEF co-cultures is shown. C Fibronectin fiber organization in HT-29 M6 co-cultured with fibroblasts. Tumor cells and the indicated MEFs were co-cultured on glass coverslips for 6 days. Co-cultures were analyzed by IF with anti-fibronectin (green) and phalloidin (white). Microscopy images are shown. D HT-29 M6 colonies co-cultured with fibroblasts control the presence fibronectin around them in an EDA-dependent manner. For each HT 29 M6 colony, the perimeter and associated empty area (black surface) were quantified (ImageJ software) from images obtained as in A. The fold-increase of the “black area/perimeter” in each co-culture with respect to values in control MEF co-cultures is shown. E The metalloproteinase inhibitor GM6001 rescues the EDA-lacking fibronectin deposition around HT-29 M6 colonies. Cocultures of HT-29 M6 and indicated MEFs were carried out and imaged as in C in the presence or absence of the 25 μM GM6001. Black area measurements and plotting were carried out as in B. Fold increase with respect to values in untreated EDA– MEF co-cultures is shown. F The metalloproteinase inhibitor GM6001 rescues the lack of fibronectin accumulation around HT-29 M6 colonies co-cultured with Snai1 KO MEFs. Co-cultures with indicated cells were established, treated and analyzed as in C and D. The fold-increase with respect to values in untreated control MEF co-cultures is shown
We also analyzed co-cultures of the colon cancer cells, HT-29 M6, which (unlike other co-cultures) grow in highly compacted colonies that are isolated from fibroblasts. In co-cultures with EDA+ MEFs, colonies of tumor cells were mostly fully encircled by fibroblasts and their fibronectin fibers. In contrast, colonies in co-cultures with EDA– MEFs were surrounded by unoccupied spaces (Fig. 5C), which were estimated with ImageJ to be eight times larger than in colonies from EDA+ MEFs co-cultures (Fig. 5D). EDA– MEFs co-cultures had broken fibronectin fibers that were clearly visualized in three-dimensional reconstructions generated from confocal images (Additional file 1: Fig. S7E), and the empty area was strongly reduced when the metalloproteinase inhibitor GM6001 was included (Fig. 5E). Altogether, these observations indicate that the activity of metalloproteinases affects fibronectin fiber organization in an EDA-dependent manner. Correlating with the fact that Snai1 KO MEFs express lower proportions of EDA+ fibronectin isoforms (Fig. 1), these MEFs also left an empty space around M6 colonies that was rescued by the MMP inhibitor when co-cultured (Fig. 5F).
Fibronectin EDA facilitates tumor cell invasionWe next took advantage that our 3D ECM can be decellularized to evaluate the activity of polymerized fibronectin fibers with or without EDA on tumor cell migration and invasion. We used MDA-MB-231 breast tumor cells, which move individually in culture (Additional file 1: Fig. S8A); we found that a higher percentage of these cells achieved oriented displacements on matrices generated by TGFβ-treated EDA+ MEFs as compared to matrices generated by TGFβ-treated EDA– MEFs or non-treated MEFs lines (Fig. 6A). Oriented movements recapitulate the alignment index of the matrices (Fig. 4G). Remarkably, an equivalent trend was obtained in invasion experiments through matrices deposited by these two TGFβ-treated MEF lines (Fig. 6B). Intermediate levels of oriented movements and invading MDA cells were obtained in matrices generated by TGFβ-treated control MEFs (Fig. 6A,B). The EDA requirement was challenged with irigenin, a small molecule that selectively binds to and blocks the EDA-integrin interaction [54]. We found that matrices generated in the presence of irigenin had a low tumor cell invasion, similar to that generated by EDA– MEFs (Fig. 6C). Irigenin treatment interfered with the orientation of the fibronectin fibers and the lacunarity of TGFβ-treated EDA+ matrices, as well with the oriented displacement of MDA cells on these matrices (Additional file 1: Fig. S8B).
Fig. 6
Tumor cell invasion is favored by the presence of EDA+ fibronectin in the 3D ECM. A MDA-MB-231 cell oriented migration depends on the presence of EDA+ fibronectin in the 3D ECM. Cell-tracker labeled MDA-MB-231 tumor cells were seeded on top of decellularized 3D ECMs generated by the indicated MEFs in the absence or presence of 5 ng/ml TGFβ1. Cell migration was recorded overnight by taking IF images every 15 min using life microscopy (Additional file 1: Fig. S8A). MDA cell movement was tracked using ImageJ software, and displacement features, such as the angle of each displacement, were measured. Oriented migration was plotted as the percentage of cell movements in the maximum orientation (up to 21˚ deviation from the mode). B MDA-MB-231 cell invasion is increased on 3D ECMs with EDA+ fibronectin. The indicated MEF lines were allowed to produce 3D ECMs in the presence of 5 ng/ml TGFβ1 on invasion-insert membranes. ECMs were decellularized, and MDA cells (in DMEM with 0.1% FBS) were seeded on top. DMEM with 10% FBS was added to the lower chamber as a chemoattractant. Cells were allowed to invade for 16 h and fixed with 4% PFA. Invading cells were stained with DAPI and quantified. C MDA-MB-231 cell invasion through EDA+ fibronectin matrices is interfered by irigenin treatment during matrix formation. MDA invasion through decellularized 3D ECMs produced by the indicated MEF lines activated with 5 ng/ml TGFβ1 and either treated or not with 50 μM irigenin, was quantified as in B. D EpRas invasion is increased on 3D ECMs containing EDA+ fibronectin. Invasion insert membranes were covered with indicated 3D ECMs as described in B and EpRas were induced to invade decellularized ECMs for 48 h and quantified as in B
We also tested EpRas cells, which formed colonies that moved collectively on decellularized matrices (Additional file 1: Fig. S8C). Of note, colonies on TGFβ-activated EDA+ matrices moved more compactly and coordinately than those on activated EDA– matrices (Additional files 2, 3, 4: S1, S2, and S3). Immunofluorescence analysis also illustrated the difference in compactness between EpRas colonies growing on each substrate (Additional file 1: Fig. S8D), and invasion assays showed that the coordinated collective movement of EpRas colonies on aligned and rigid fibronectin EDA+ matrices was accompanied by a higher capacity to invade the matrix (Fig. 6D).
Matrix remodeling by TGFβ is required to stimulate fibroblastFibroblasts are activated by recombinant EDA fragments [31]. Here, we tested whether EDA in extracellular-deposited fibronectin fibers was also effective. Naïve fibroblasts were seeded on decellularized 3D ECMs, and the presence of cytosolic α-SMA stress fibers (a marker of fibroblast activation) was analyzed 16 h later. For both mouse mesenchymal stem cells (MSC) and NIH3T3 fibroblasts, the percentage of cells with α-SMA–positive fibers was higher on matrices derived from EDA+ MEFs than EDA– MEFs (Fig. 7A; Additional file 1: Fig. S9).
Fig. 7
Matrices with EDA+ fibronectin induce the assemblage of α-SMA fibers in naïve fibroblasts. A MSC or NIH3T3 fibroblasts are induced to assemble α-SMA fibers by EDA+ matrices. MSC or NIH3T3 were grown 24 h on decellularized 3D ECMs generated by EDA– or EDA+ MEFs before detecting α-SMA and nuclei by IF. Images obtained through fluorescence microscopy were used to quantify the percentage of cells presenting α-SMA positive stress fibers. B NIH3T3 fibroblasts are induced to assemble α-SMA fibers by control and EDA matrices. NIH3T3 fibroblasts were grown 24 h on decellularized 3D ECMs generated by the indicated MEFs in the presence or absence of 5 ng/mL TGFβ1. The percentage of fibroblasts presenting α-SMA positive stress fibers was quantified as in A. C Irigenin interferes with fibroblast activation by EDA+ matrices. NIH3T3 fibroblasts were grown for 24 h on decellularized 3D ECMs generated by the indicated MEFs in the presence or absence of 5 ng/mL TGFβ1, and the presence or absence of 50 μM irigenin. The percentage of fibroblasts presenting α-SMA positive stress fibers was quantified as in A
Matrices deposited by MEF EDA+ in either the presence or absence of TGFβ stimulated NIH3T3 (Fig. 7B); however, matrices deposited by control MEFs stimulated fibroblasts only if generated in the presence of the TGFβ cytokine (Fig. 7B). Given the elevated EDA+ fibronectin ratio generated and deposited by control MEFs (Figs. 1 and 4), our result suggests a TGFβ-induced conformational change that exposes polymerized EDA to fibroblasts.
To validate that stimulation of naïve fibroblasts depends on EDA in our experimental approach, fibroblast stimulation was challenged with irigenin. Addition of this compound prevented the formation
Comments (0)