
記住我
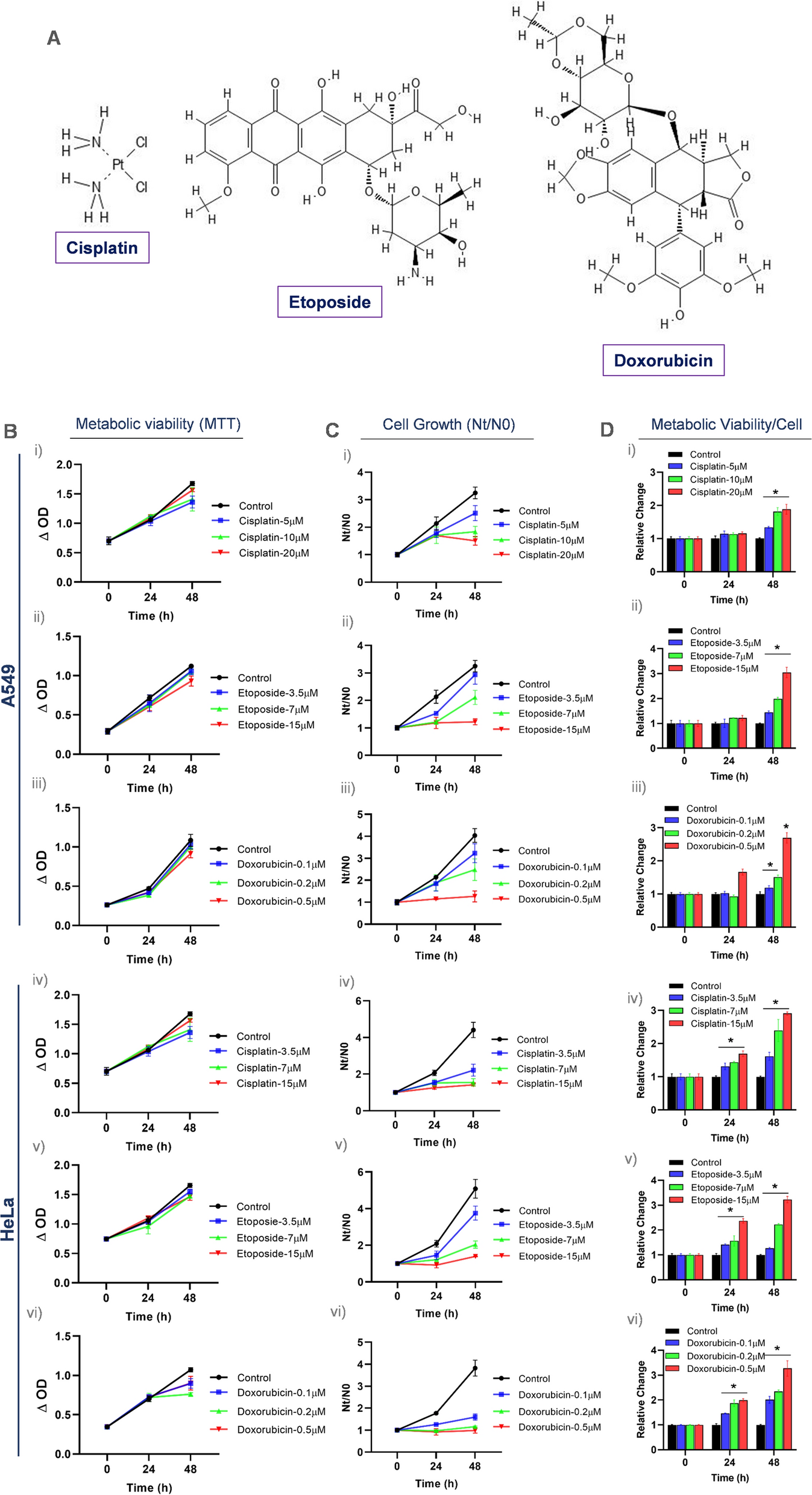
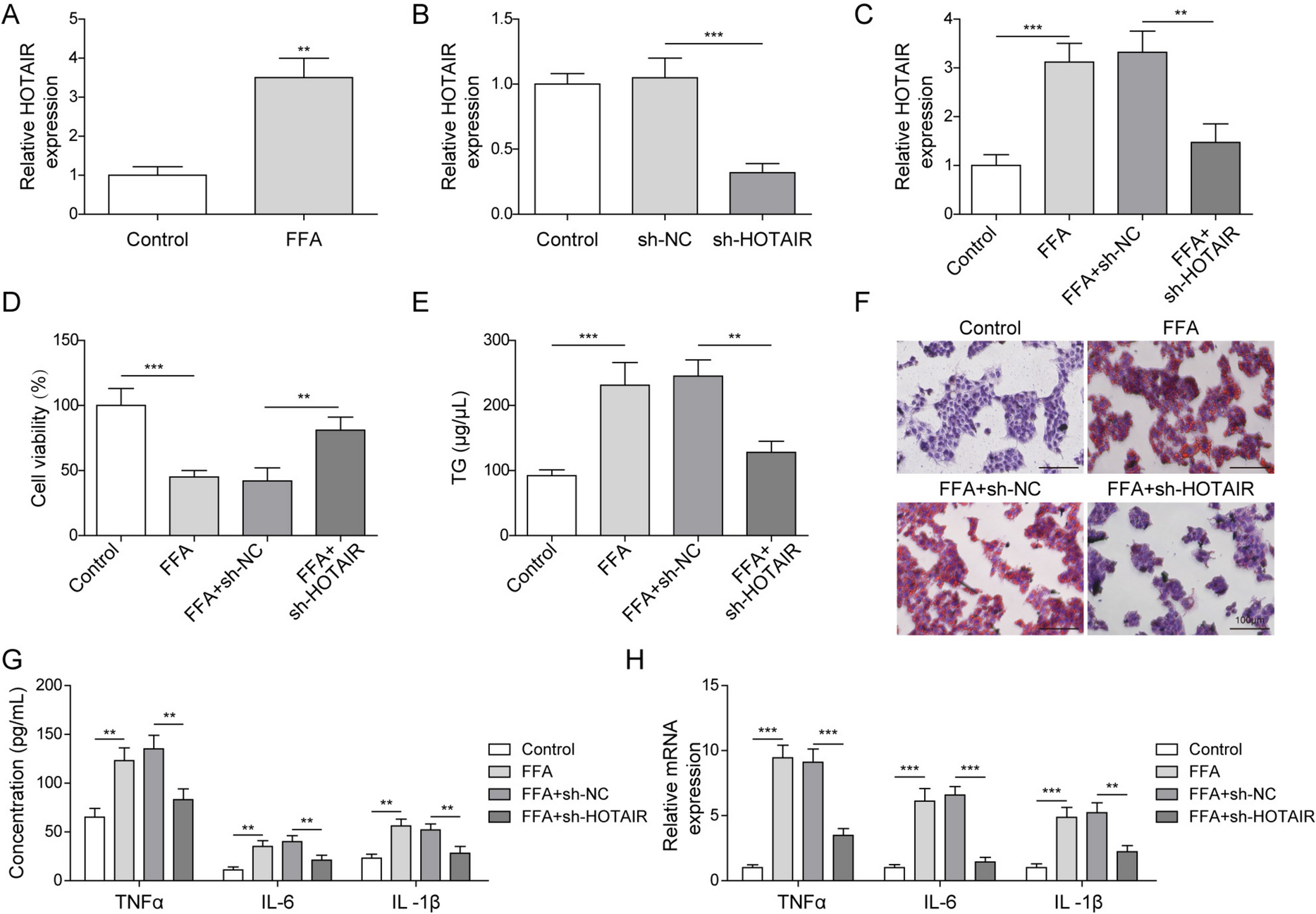
A patented CVA6-VLP-producing CHO-S cell line (Kuwabara et al. 2020) provided by BIKEN Group Japan (Osaka, Japan) was cultivated in CD FortiCHO™ medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 8 mM glutamine (Fujifilm Wako, Osaka, Japan). Cell cultures using 0.5 L Optimum Growth® Flasks (Thomson Instrument, CA, USA) with 0.1 L working volume were performed after cell thawing and expansion. Cell cultivation was performed in a humidified incubator (Climo-Shaker, Kuhner, Switzerland) operated at 37 °C, 8% CO2, and 140 rpm. Cells were subcultured every 3–4 days when they were during the exponential phase prior to fed-batch bioreactor cultivation.
Fed-batch laboratory-scale bioreactor cultivationLaboratory-scale bioreactor cell culture was performed using a stirred tank glass bioreactor (ABLE Biott, Tokyo, Japan) with a 2 L maximum working volume operated in fed-batch mode. Inoculation was performed by seeding cells at 4 × 108 cells/L in a 0.8 L working volume. Cultivation conditions were controlled at 30% dissolved oxygen using a pure O2 sparge, 80 rpm agitation, 0.1 L/min constant air flow, and 8 × 10–3 L/min constant CO2 flow. Temperature was controlled at 37 °C and then changed to 32 °C after day 5. pH maintenance was conducted by sparging CO2 gas and 1 M NaHCO3 addition. In the control experiment (bioreactor 1), pH was maintained at 7.15 until the end of culture. In the treatment experiment (bioreactor 2), pH was maintained at 7.15 and then changed to 6.75 after day 10.
Up to 3 × 10–3 L FoamAway™ (Thermo Fisher Scientific) was added to when foaming was observed during cell culture. From day 3, 1.6 × 10–2 L of feed medium (EfficientFeed™ C + AGT™ Supplement, Thermo Fisher Scientific), which was equal to 2% of the initial volume, was fed into the bioreactor every day. Additional glucose supplementation was conducted by adding a 300 g/L glucose stock solution to ensure a glucose concentration of > 2 g/L. Then, 4 × 10–3 L of sample was collected from the bioreactor daily for cell growth, CVA6 VLP concentration, and metabolite analyses.
Cell counting and metabolite measurementsViable cell concentration and viability were measured by a ViCell automated cell counter (Beckman Coulter, Brea, CA, USA). Metabolite (glucose, glutamine, lactate, and ammonia) concentrations were quantified using a BioProfile 400 Automated Chemistry Analyzer (Nova Biomedical, Waltham, MA, USA).
Cell lysis quantificationCell lysis quantification was performed by adapting a previously proposed method that uses double-stranded DNA (dsDNA) as an indicator (Klein et al. 2015). Briefly, dsDNA in supernatant samples was measured using a Quant-iT PicoGreen dsDNA Assay Kit (Thermo Fisher Scientific). Supernatant samples were diluted appropriately and 1 × 105 μg/L Lambda DNA Standard was diluted to prepare the standard curve. Samples and the standard were mixed with assay reagent in a 96-well plate. After incubation at room temperature for 2 min, a VICTOR Nivo Multimode Microplate Reader (PerkinElmer, Waltham, MA, USA) was used to excite samples and the standard at 492 nm, and fluorescence emission intensities at 530 nm were measured. Intracellular dsDNA content was determined by the mass balance under the assumption that cell growth stopped after the exponential phase (after day 5). By dividing dsDNA in the supernatant by intracellular dsDNA content, the lysed cell density was quantified. The spontaneous release of dsDNA from viable cells was neglected because dsDNA concentration in the supernatant during the exponential phase was negligible compared with that during the dead phase.
CVA6-VLP quantificationA sandwich enzyme-linked immunosorbent assay developed by Biken Group Japan was applied for CVA6 VLP quantification. Briefly, 96-well plates were coated with an anti-CVA6 monoclonal antibody and incubated at 4 °C overnight. Non-specific binding was blocked by incubation with EzBlock Chemi (Atto, Tokyo, Japan) blocking buffer for 1 h at 37 °C. Then, microplates were washed thrice with washing buffer (0.05% (v/v) Tween 20 in phosphate-buffered saline) prior to addition of the standard or samples. Microplates were incubated at 37 °C for 1 h, followed by three washes. A horseradish peroxidase-conjugated anti-CVA6 monoclonal antibody was added as the secondary antibody. After 1 h of incubation at 37 °C, microplates were washed three times and binding was visualized by incubation with a TMB solution (Surmodics, MN, USA) for 15 min. To stop the reaction, BioFX 650 nm Liquid Stop Solution for TMB Microwell substrates (Surmodics) was added to each well. Absorbance at 650 nm was read by a microplate reader (Corona Electrics, Ibaraki, Japan).
Model constructionKinetics between cells, major metabolites, and VLP were described by ordinary differential equations. Volume changes during fed-batch cultivation were disregarded because they were negligible compared with the working volume.
$$r = r_ \cdot \left( }} }}} \right)$$
(1-b)
where \(_\) denotes total cell density; \(r\) denotes intrinsic growth rate and \(_\) denotes maximum intrinsic growth rate; \(_\) denotes maximum total cell density.
A logistic equation (Martínez et al. 2020; Shirsat et al. 2015a, b) was applied to describe the behavior of the total cell density (\(_\)) to prevent overfitting because it had fewer parameters than Monod-type equations. This equation also complied with the previous assumption that cell proliferation stopped during the late stage of culture.
$$\frac }} = k_ X_ - k_ X_$$
(2-a)
If (\(_\)>\(_\) during dead phase):
$$k_ = k_ \cdot \frac }} + c_ - c_ }} }} }}$$
(2-b)
Else:
$$\frac }} = k_ X_ + k_ X_$$
(3)
$$\frac }} = rX_ - \left( + k_ } \right) \cdot X_$$
(4)
where \(_\) is dead cell density and \(_\) is viable cell density; \(_\) is specific death rate and \(_\) is specific lysis rate from dead cells; \(_\) stands for death rate during dead phase; \(__}\) stands for constant for cell death due to ammonia accumulation, \(_\) stands for ammonia concentration and \(_\) stands for critical ammonia concentration for specific death rate; \(_\) is minimum death rate and \(_\) is specific lysis rate from viable cells; \(_\) denotes lysed cell density.
Cell death and lysis pathways as well as their corresponding coefficients are illustrated in Fig. 1. These pathways were modeled by adapting previously proposed equations (Kontoravdi et al. 2007; Kroll et al. 2017). Briefly, cell death was accelerated significantly when the ammonia concentration exceeded its critical concentration (\(_\) = 5 mM) during the dead phase (Kontoravdi et al. 2007). Additionally, lysed cells (\(_\)) could be generated by viable cells (\(_\)) directly or dead cells (\(_\)) as shown by the two terms in Eq. (3), respectively. \(_\) was the difference between \(_\) and the sum of \(_\) and \(_\).
$$\frac }} = - \left( - \left( + k_ } \right)X_ } \right)/Y_ /Glc}} - m_ X_ + \frac c_ }} + \frac c_ }}$$
(5)
Fig. 1
Adapted from Kroll et al. (2017). This figure was created by Adobe Illustrator 2023 (Adobe Inc., CA, USA)
Cell death and lysis pathways with their corresponding rate coefficients.
Before day 5:
$$\frac }} = \left( - \left( + k_ } \right) \cdot X_ } \right)/Y_ /Glc}} - m_ X_ } \right) \cdot Y_$$
(6-a)
After day 5:
$$\frac }} = \left( - \left( + k_ } \right) \cdot X_ } \right)/Y_ /Glc}} - m_ X_ } \right) \cdot Y_ - q_ X_$$
(6-b)
$$q_ = q_ \cdot \frac }} + c_ }}$$
(6-c)
where \(_\) and \(_\) are glucose and lactate concentrations, respectively; \(__/Glc}\) is yield coefficient of cell proliferation to glucose uptake; \(_\) and \(_\) denote the volumes of feed medium and glucose supplementation at \(i\) th feeding, respectively; \(_\) and \(_\) are the glucose concentrations of the feed medium and glucose supplementation, and \(V\) is the working volume; \(_\) is specific glucose consumption rate for cell maintenance; \(_\) represents yield coefficient of lactate production to glucose uptake and \(_\) represents specific lactate uptake rate; \(_\) is maximum lactate uptake rate and \(_\) is Monod kinetic constant for lactate uptake.
Equation (5) was modified from a study by Xing et al. which consists of glucose consumption due to cell growth and maintenance as well as the increase in glucose concentration as a result of feeding (Xing et al. 2010). The change of lactate concentration was modeled by Eq. (6). Cells switched from lactate production to lactate consumption after day 5, which can be observed in Fig. 2i. The mechanism of this kind of lactate switch remains unclear, although it is common in CHO cell lines (Hartley et al. 2018). A yield coefficient (\(_\)) was used to link glucose use and lactate production. After day 5, an additional Monod-type term was applied to account for lactate consumption.
Fig. 2
This figure was created by MATLAB R2021a (The Math Works, Inc., MA, USA)
Comparison between model simulations of bioreactor 1 (solid line) and bioreactor 2 (dash line) (Edward 2022) versus experimental data of bioreactor 1 (closed circle) and bioreactor 2 (open circle) for nine variables (a–i). R2 was used to evaluate the goodness of fit.
If (\(_\) > 0 mM):
$$\frac }} = - \left( - \left( + k_ } \right) \cdot X_ } \right)/Y_ /Gln}}$$
(7-a)
$$\frac }} = \left( - \left( + k_ } \right) \cdot X_ } \right)/Y_ /G\ln }} \cdot Y_$$
(8-a)
If (\(_\) = 0 mM):
$$\frac }} = - \left( - \left( + k_ } \right) \cdot X_ } \right)/Y_ /Amm}} + q_ X_$$
(8-b)
where \(_\) and \(_\) denote glutamine and ammonia concentrations, respectively; \(__/Gln}\) denotes yield coefficient of cell proliferation to glutamine uptake and \(_\) denotes yield coefficient of ammonia production to glutamine uptake; \(__/Amm}\) yield coefficient of cell proliferation to ammonia uptake and \(_\) is specific ammonia production rate.
Similarly, glutamine consumption was described by a yield coefficient and ammonia was considered to be the production of glutamine when glutamine was available. When glutamine was depleted, ammonia was used as an alternative nitrogen source and consumed until the end of the cell growth phase. This phenomenon has also been reported in other mammalian cell lines (Lie et al. 2019). Additionally, ammonia production for cell maintenance was modeled through \(_\), which differs at various pHs. After day 6. If (\(_<_\) and pH = 7.15):
If (\(_=_\) or pH = 6.75):
$$\frac }} = VLP_ \cdot \left( + k_ } \right) \cdot X_$$
(10)
where \(_\) is the amount of VLPs accumulated intracellularly, and \(_\) is its accumulation rate; \(_\) denotes VLP concentration in the medium.
Our previous observations indicated that the VLPs were produced efficiently after day 6. Moreover, a pH downshift suppressed production of intracellular VLPs. Additionally, there was a maximal intracellular VLP content that a single cell can contain (\(_\)). The VLP concentration in the medium was simulated by combining \(_\) with cell release, either cell death or lysis, as shown by Eq. (10).
The computational work was conducted in MATLAB R2021a and Optimization Toolbox (The Math Works, Inc., MA, USA). Ordinary differential equations were solved by ode45 solver in MATLAB. Parameters were determined by minimizing weighted root-mean-square deviation between the experimental data and model simulation. Model simulation was evaluated by the coefficient of determination (R2) (Colin Cameron and Windmeijer 1997).
Local sensitivity analysisTo further understand the effect of various model coefficients and initial culture conditions, local sensitivity analysis was carried out. Briefly, the relative change in the final VLP concentration by the end of cell culture was simulated in response to a 10% change in model parameters independently. Model parameters of bioreactors 1 and 2 before day 10 were averaged because two bioreactors were operated under the same conditions before the pH shift on day 10. Coefficients after the pH shift were based on the simulation of bioreactor 2. Local sensitivity was simulated under the same pH shift strategy in bioreactor 2.
Predictions under various pH shift timingsAfter model calibration, determined parameters were used to predict the experimental performance of the VLP production process under various pH shift strategies. Various time-dependent cell densities, major metabolite concentrations, and VLP yield were predicted to understand and optimize the process of VLP vaccine production.
Multiple objective optimizationDesirability methodology was applied to optimize the production process with multiple responses (\(_\)) such as metabolite concentrations or the VLP concentration. In brief, various responses were used to compute an individual desirability function (\(_\left(_\right)\)), which was either maximization (Eq. 11-a) or minimization (Eq. 11-b) depending on customized objectives (Möller et al. 2019). Individual desirability \(_\left(_\right)\) ranged from 0 to 1. The objective was satisfied if its \(_\left(_\right)\) was close to 1. The overall desirability was obtained by multiplication of individual objectives as illustrated by Eq. (12), which was used as a quantitative criterion for comparison to guide decision making.
$$d_ \left( } \right) = \left\l} 0 \hfill & } y_ < L_ } \hfill \\ - L_ }} - L_ }}} \right)} \hfill & } L_ < y_ < U_ } \hfill \\ 1 \hfill & } y_ < U_ } \hfill \\ \end } \right.$$
(11-a)
$$d_ \left( } \right) = \left\c} }\,y_ < L_ } \\ - y_ }} - L_ }}} \right)\quad }\,L_ < y_ < U_ } \\ }\,y_ > U_ } \\ \end } \right.$$
(11-b)
$$D = \mathop \prod \limits_} = 1}}^}} d_ \left( } \right) = d_ \left( } \right) \times d_ \left( } \right) \cdots \times d_ \left( } \right)$$
(12)
where \(_\) and \(_\) are customized upper and lower boundaries, respectively. By integrating the designed desirability function with the model simulation, multiple objective optimization was performed to predict the optimal pH shift strategy and timing to stop the batch.
留言 (0)