
Remember me
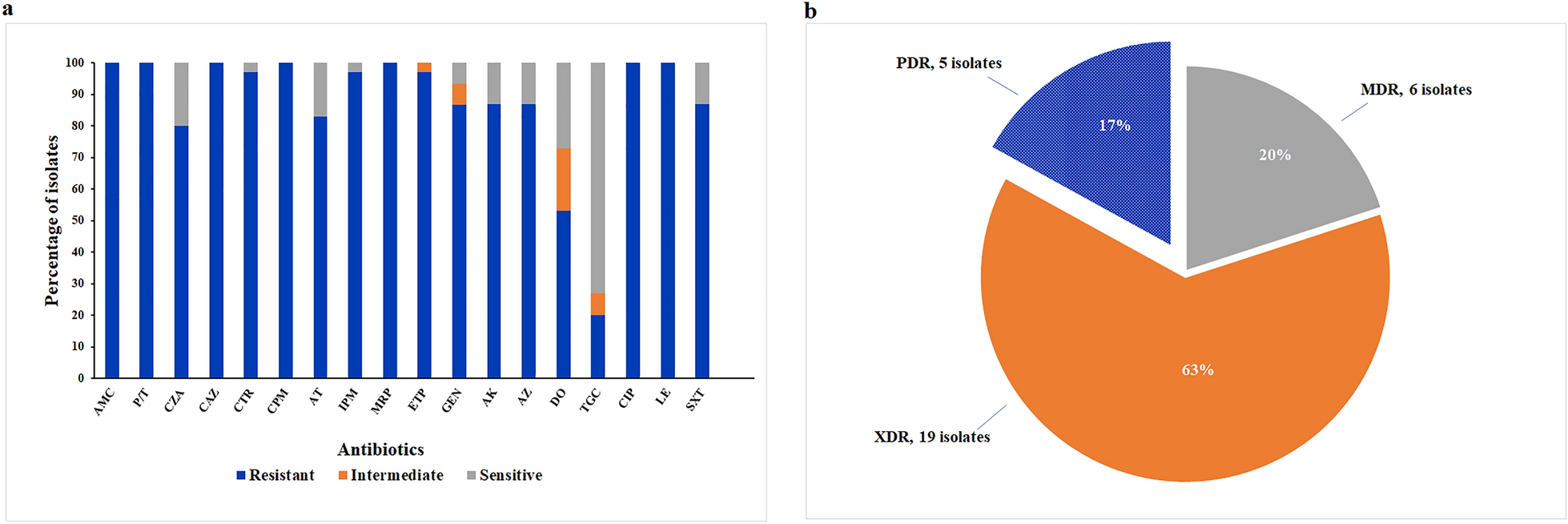



Total of 69 subjects, including 30 control subjects (S-C) and 39 patients with cirrhosis (S-LC), with similar demographics, were included in this study and the information were shown in Tables 1 and 2. We compared the alpha diversity of gastric microbiota between the S-LC and S-C group, and the Sobs (P < 0.05), Shannon (P < 0.001), Ace (P < 0.05), and Chao1 (P < 0.05) values were significantly higher in S-LC (Additional file 1: Table S2, Fig. 1A). Meanwhile, beta diversity analysis showed separate clusters for S-LC and S-C (P = 0.001, Fig. 1B). The gastric microbiota was dominated by eight phyla: Proteobacteria, Campilobacterota, Firmicutes, Bacteroidetes, Actinobacteria, unclassified_k__norank_d__Bacteria, Fusobacteria, and Cyanobacteria (Fig. 1C), although the two groups presented in different order of relative abundance at the phylum level. The gastric microbiota in liver cirrhosis had an over-representation of Firmicutes, Bacteroidetes, Actinobacteria, and Fusobacteriota (P < 0.001; Additional file 3: Figure S2). At the genus level, several genera, including Streptococcus, Prevotella, Neisseria, Fusobacterium, Haemophilus, Veillonella, Porphyromonas, Actinomyces, Gemella, Alloprevotella, Rothia, Granulicatella, and Peptostreptococcus, significantly increased in relative abundance between the S-C and S-LC (Fig. 1D). The Helicobacter and Achromobacter decreased in LC (Fig. 1D).
Fig. 1
Gastric mucosal microbiome dysbiosis between S-C and S-LC. S-C represents gastric mucosa derived from the control group, and S-LC represents gastric mucosa derived from the liver cirrhosis group. In A, B, D–F, red is the S-C group, and blue is the S-LC group, *p < 0.05, **p < 0.01, ***p < 0.001. A Increased microbial richness, estimated by Shannon index. B Gastric mucosal microbiota showed relative clustering between control subjects compared with all patients with cirrhosis. C Relative abundance of microbial species at the phylum level. D The difference in species composition at the genus level. E The LDA value distribution histogram shows the species between S-C and S-LC at the genus level (LDA > 4). F The relative abundance of H. pylori was higher in S-C than in S-LC. Statistical significance was determined by the Mann–Whitney U test
Bacteria differentially abundant in LC versus controlsWe performed linear discriminant analysis effect size at the genus level to further identify the gastric-specific species signatures. Seven bacterial taxa showed distinct relative abundances between the two groups. Increased abundance in bacteria, including Streptococcus, Neisseria, Prevotella, Veillonella, and Porphyromonas, and decreased abundance in Helicobacter and Achromobacter were observed in S-LC (LDA score > 4, P < 0.05; Fig. 1E). Furthermore, we found that the H. pylori infection was significantly lower in S-LC than in S-C (Fig. 1F).
To understand the potential interplay among differentially abundant bacteria in S-LC and control, we performed the network topology analysis (Additional file 4: Figure S3) at the genus level. In S-C group, co-occurrence interactions were observed, reflecting synergistic interkingdom interactions’ contribution to gastric microbiota homeostasis (Additional file 4: Figure S3A). Much fewer co-occurrence interactions were observed in S-LC (Additional file 4: Figure S3B). In addition, H. pylori had a co-exclusive association with other gastric microbes in S-C predominantly. However, these correlations were relatively infrequent in S-LC. Collectively, the above microbial analysis indicated a state of dysbiosis in the mucosal microbiome of LC patients.
Blood metabolism changes with LCWe recruited 22 cirrhotic patients (age 53.04 ± 11.08 years, 15 men) and 44 control patients (age 48.32 ± 14.37 years, 25 men) who agreed to give blood serum (Tables 1, 2). We used liquid chromatography-mass spectrometry; to analyze the serum samples, and the abundance profiles were obtained for 1540 annotated serum metabolites. It was found that 492 of 1540 metabolites had significantly different abundances (Fig. 2A). We subsequently performed the partial least squares discriminate analysis and the results revealed visual separation between these groups without overfitting (Fig. 2B). In addition, PLSDA-VIP table (Fig. 2C, Additional file 1: Table S3) showed top 30 metabolites with VIP > 1.0 and P < 0.001. Of them, 9 were increased in the control group, and 21 were increased in the LC group. Notably, the VIP value of taurochenodeoxycholate-3-sulfate (VIP = 3.72), taurodeoxycholic acid (VIP = 2.93), and cis-5-tetradecenoylcarnitine (VIP = 2.89) were more than 2, indicating their significant contribution to the disease. Furthermore, we carried out an enrichment analysis for all differential metabolites using the KEGG. The results indicated that the metabolites in LC were mainly associated with sphingolipid metabolism, glycerophospholipid metabolism, cysteine and methionine metabolism (Additional file 5: Figure S4).
Fig. 2
The liver cirrhosis group is associated with altered serum metabolites. A Differential metabolites by volcano diagram. B PLS-DA of the metabolites across the two groups. C VIP scores with the corresponding expression heatmap. On the left side is the metabolite heatmap. On the right side is the metabolite VIP bar graph. The bar length indicates the contribution of the metabolite to the difference between the two groups. The higher value means the metabolite is more difference between the two groups. The bar color indicates the P value of the metabolite between the two groups, *p < 0.05, **p < 0.01, and ***p < 0.001
Association of microbes and metabolites with LCTo further determine the relationships between gastric microbiota and metabolic changes, we subsequently perform the Spearman’s correlation analysis of serum differential metabolite and microbiota in stomach. Interestingly, there was a strong correlation between a large number of stomach microbiota and altered metabolites (Fig. 3A). The correlation was considered statistically significant with correlation coefficient |r|> 0.7 and p < 0.05 in this study, and results were visualized as heatmaps.
Fig. 3
Correlation analysis between the microbiota and metabolites. A Heatmap of gastric mucosal flora and serum metabolite correlation. Asterisk(***) denotes a P value less than 0.05 and |r|> 0.7. B Heatmap of gastric mucosal flora and serum long-chain ACs correlation. Asterisk(***) denotes a P value less than 0.05 and |r|> 0.6. C Heatmap of gastric mucosal flora and serum bile acids correlation. Asterisk(***) denotes a P value less than 0.05 and |r|> 0.7
It was found that genus Streptococcus, enhanced in the gastric microbiota community in LC patients, was positively correlated with taurodeoxycholic acid (r = 0.71) and (E)-2-Penten-1-ol (r = 0.82). In addition, His Phe Trp and phosphatidylserine were negatively correlated with almost all the bacteria except Helicobacter (r = 0.65; r = 0.69). The relative concentrations of some long-chain acylcarnitines (palmitoyl-l-carnitine, octadecenoylcarnitine, 3-hydroxyhexadecanoyl carnitine, gamma-linolenic carnitine, and cis-5-tetradecenoylcarnitine) were generally seen to increase in the serum metabolites from cirrhotic patients. As shown in Fig. 3B, increased long-chain ACs were positively associated with almost all the bacteria, such as Actinomyces and cis-5-tetradecenoylcarnitine (r = 0.66). Besides, taurodeoxycholic acid, deoxycholic acid glycine conjugate, glycocholic acid, taurocholic acid, cholic acid, taurochenodeoxycholate-3-sulfate, taurochenodeoxycholate-7-sulfate, chenodeoxycholic acid 3-sulfate, and their derivatives were positively correlated with almost all the bacteria, except 7a,12a-dihydroxy-3-oxo-4-cholenoic acid (Fig. 3C).
Gastric mucosal transcriptome changes in LCTo obtain a comprehensive view of gastric mucosa influenced by microbial colonization and metabolic alterations, we further investigated the gastric mucosal transcriptome in two groups comprising 10 control and 10 LC patients. It was found that 181 and 124 genes were differentially down and upregulated, respectively. Of them, the endothelial cell specific molecule 1, serpin family E member 1, mucin 2, caudal type homeobox 2, and retinol binding protein 2 were significantly upregulated genes (Fig. 4A, B) and were associated with intestinal metastasis and gastric carcinogenesis [15,16,17,18]. We also found that defensin alpha 5 (DEFA5), a gene encoding an antimicrobial peptide, is significantly upregulated (P < 0.001). Then, we mapped all DEGs to KEGG pathways, and the top 11 specific pathways were represented in a bubble chart (Fig. 4C). Furthermore, genes associated with the most significantly enriched pathways (q < 0.05) were shown in Additional file 6: Figure S5. Among these 11 pathways, “neuroactive ligand-receptor interaction” was the most represented pathway. Significantly, it was found that the bile secretion signaling pathway was significantly upregulated in the LC group. In the bile secretion signaling pathway, solute carrier family 51 subunit alpha (SLC51A), solute carrier family 51 subunit beta(SLC51B), cytochrome P450 3A4(CYP3A4) were up-regulated.
Fig. 4
S-C represents gastric mucosa derived from the control group, and S-LC represents gastric mucosa derived from the treatment group. A Volcano map of DEGs. B Venn graph of DEGs. C DEGs enriched in the KEGG pathway. The X-axis represents the rich factor, indicating the ratio of enriched genes to total genes in this pathway. A more prominent rich factor indicates more significant enrichment. ESM1 endothelial cell specific molecule 1, SERPINE1 serpin family E member 1, MUC2 mucin 2, CDX2 caudal type homeobox 2, RBP2 retinol binding protein 2, DEFA5 defensin alpha 5, DEG differential expression gene
Comments (0)