
Remember me
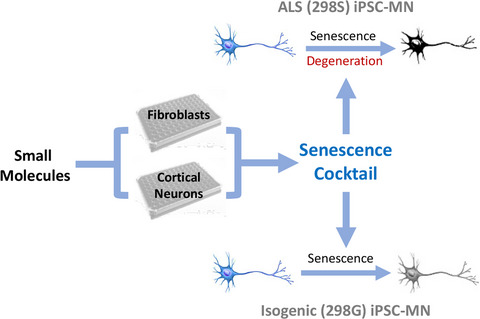
As global life expectancy increases, neurodegenerative disorders are predicted to cause a staggering burden to society. Substantial efforts have been made to develop effective therapies, but progress is slow and drugs developed based on animal studies have so far mostly failed in clinical trials (Mitsumoto et al., 2014; Paganoni et al., 2014). Poor clinical translatability of animal models necessitates additional models to test therapeutic strategies.
Human pluripotent stem cell (hPSC)-derived neurons model early stages of neurodegeneration and have potential benefits in drug discovery and testing. An advantage of the hPSC model is the ability to capture the human genetic background underlying diseases by establishing patient-specific iPSCs, or by studying specific effects of disease proteins via introducing disease-related mutations into otherwise normal hPSCs (Chen et al., 2014; Sances et al., 2016). However, the iPSC reprogramming process erases many of the aging marks found in somatic donor cells (Lapasset et al., 2011; Menendez et al., 2011; Miller et al., 2013; Rando & Chang, 2012), and hPSC-derived neurons are similar to those in fetal development, based on transcriptional and functional profiling (Nicholas et al., 2013; Patterson et al., 2012; Vera & Studer, 2015). Thus, generating hPSC-derived neurons that resemble those in the adult and aging brain is critical for modeling neurodegenerative diseases using hPSCs.
One approach for modeling cellular senescence (CS) is trans-differentiation of fibroblasts or other aged somatic cells into neurons, avoiding the pluripotent stage and maintaining senescence markers (Mertens et al., 2015; Vierbuchen et al., 2010). Indeed, a recent study showed that transdifferentiated neurons retain age-related transcription profiles and manipulation of RANBP17 was able to reverse some of the age-related transcriptional changes in iPSC-derived neurons (Mertens et al., 2015). However, direct conversion of fibroblasts into neurons is relatively low throughput given the lack of expansion capacity of the resulting neurons.
Modulation of genes linked to premature aging disorders is another strategy to accelerate aging in stem cell models. The ectopic expression of progerin, a mutant form of nuclear lamina protein A (LMNA) that causes accelerated aging in progeria, in an iPSC model of Parkinson's disease (PD) can trigger age-related and degenerative phenotypes, including neuromelanin accumulation, dendrite degeneration, loss of tyrosine hydroxylase, and accumulation of pathological aggregates (Miller et al., 2013). It remains to be determined how closely these approaches model physiological aging in normal neurons or pathological aging seen in late-onset diseases. Overexpression of premature aging genes introduces the challenge of distinguishing phenotypes related to the disease from those induced by foreign gene overexpression.
In the present study, we screened for chemicals/pathways that selectively trigger senescence phenotypes in primary neonatal fibroblasts and iPSC-derived cortical neurons. To identify pathways important in neuronal senescence, we first used transdifferentiated neurons from aged and young fibroblasts and identified molecular markers for neuronal aging, including decreased expression of H3K9Me3, chromatin-associated protein HP1γ, and lamina-associated polypeptide Lap2β. We then used these readouts for screening small molecules and developed a combination of molecules that induce senescence and protein aggregation in cortical neurons differentiated from hPSCs. We evaluated this chemical-induced senescence (CIS) approach in motor neurons (MNs) derived from ALS (TARDBP mutant) patient iPSCs and confirmed that CIS promoted earlier and consistent manifestation of disease-related phenotypes. Furthermore, using autophagy activator molecules, we were able to mitigate CS phenotypes in the MNs. Thus, this CIS strategy enables more effective iPSC modeling of phenotypes in ALS.
2 RESULTS 2.1 Identification of small molecules that induce senescence in neonatal fibroblastsPrimary human fibroblasts retain age-related markers depending on the age of the individual from which the cells are isolated (Childs et al., 2015). These cells are thus appropriate reference for studying CS. We compared neonatal fibroblasts with those from a 72-year-old male and 62-year-old female donors by examining the expression of age-related markers H3K9Me3, Lap2β, and HP1γ. We found that neonatal fibroblasts expressed a higher level of H3K9Me3, Lap2β, and HP1γ than old fibroblasts (72 years) in our high-content imaging platform. In addition, the old fibroblasts expressed the senescence-associated β-Gal (Figures 1a,b and S1A-C). These findings are consistent with a previous observation (Miller et al., 2013), indicating that these markers are reliable readouts for assessing CS.

Identifying small molecules for inducing CS in human neonatal fibroblasts. (a) Immunostaining for H3k9Me3, Lap2β, and HP1γ proteins in both neonatal and aged fibroblasts. Scale bar = 100 μm. (b) Frequency distribution for different bins of signal intensity in high-content imaging for H3k9Me3, Lap2β, and HP1γ proteins in male neonatal and aged (72 years old) fibroblasts. (c) Frequency distribution for H3k9Me3, Lap2β, and HP1γ protein expression in male neonatal fibroblasts treated with different small molecules; dashed red line is control, and top seven molecules for each protein are shown in the graph. (d) Mean difference in signal intensity for all 25 small molecules depicted as mean ± 95% confidence intervals compared to the DMSO control group. The zero line means no difference compared to control and if the difference does not touch the reference line then the changes in expression are significant (n = 4, t test compared to the control DMSO)
We then looked for molecules that may induce senescence phenotypes in the neonatal fibroblasts, focusing on the known senescence associated pathways. We selected 25 small molecules known to affect pathways involved in CS (Petrova et al., 2016), including autophagy-related molecules, Akt signaling, and inhibitors of mTOR, HDAC, ZMPSTE24, and Sirtuin signaling (Table S1). We examined the toxicity of these molecules in their minimum effective concentrations based on previous studies using calcein AM and ethidium homodimer (EthD-1) fluorescent dyes to distinguish live versus dead cells. None of the small molecules induced cell death beyond the DMSO control (5%–10% cell death) at the final selected concentration (Figure S1D). By culturing the neonatal fibroblasts in the presence of the small molecules at an effective dose for five consecutive days and examining the expression of the above CS markers, we found that more than half of the molecules (13 molecules, p ≤ 0.001, Table S2) significantly decreased the expression of all three readouts (Figure 1c,d). Among the 13 molecules, seven also induced expression of β-Gal, another consensus marker for CS (Figure S1E,F). Thus, we identified a set of small molecules that induce senescence phenotypes in neonatal fibroblasts.
2.2 Identifying small molecule cocktails that enhance neuronal senescenceEpigenetic marks, including those associated with aging, are largely erased during reprogramming to iPSCs (Lo Sardo et al., 2017; Rando & Chang, 2012). Consequently, cells differentiated from iPSCs, including neurons, behave like those in embryonic development. In contrast, neurons directly converted from fibroblasts by forced expression of transcription factors retain much of the age-related signatures in their parental somatic cells (Mertens et al., 2015). To validate this phenomenon and to establish CS readouts in neurons, we reprogrammed both young and old human fibroblasts to neurons using a combination of gene overexpression and small molecules (Mertens et al., 2015). Both neonatal and aged fibroblasts were transduced with lentiviral particles for Eto and XTP-Ngn2:2A:Ascl1 (N2A) and cultured in the presence of G418 and puromycin for at least three passages. Induced neurons (iNs), exhibiting polarized morphology and expressing neuronal proteins like β-III tubulin (Figure 2a), appeared at the 2nd week in the neonatal fibroblast group and mostly at the 3rd week for the old fibroblast group. At the end of 3 weeks of DOX treatment, the mean conversion rate for neonatal iNs was 18.1% ± 3.5, whereas for aged iNs was 39% ± 4.4, which were used for further experiments without any purification (Figure 2b). Importantly, the iNs from old fibroblasts showed a lower intensity in the epigenetic mark H3K9Me3, Lamin B2, and Lap2β as well as the heterochromatin protein HP1γ (Figure 2c). Besides the above markers, the morphology and size of a cell and nucleus may serve as a sign of CS (Zhao & Darzynkiewicz, 2013). We also noticed that neonatal iNs had a lower Hoechst (nuclear) intensity (Figure 2d) and a smaller nucleus area compared to their aged counterparts (Figure 2e), while there were no differences in the nuclear roundness and ratio between young iNs and aged iNs (Figure 2f,g). Our results confirmed that the iNs from aged fibroblasts retain the age-related signatures of their parental cells, setting a reference for us to examine the effects of small molecules on CS in embryonic neurons.

Cellular senescence marks are preserved during direct reprograming of fibroblasts to neurons. (a) Immunostaining for H3K9Me3, LaminB2, Lap2β, and HP1γ co-stained with TUJ1 (red) in induced neurons (iNs) derived from fibroblasts of neonatal and a 72-year-old donor. Scale bar = 100 μm. (b) Percentage of TUJ1-positive neurons. (c) Mean signal intensity for H3K9Me3, Lap2β, LaminB2, and HP1γ. (d) Hoechst signal intensity, (e) nucleus roundness, (f) nucleus ratio, and (g) nucleus area for both young and aged iNs (n = 3–10, ns: not significant, *p < 0.05, **p < 0.01, ***p < 0.001 unpaired t test). Note that the representative images of Hoechst and Tuj1 staining for young and aged iNs are placed after D-G
Neurons differentiated from ESCs and iPSCs resemble those during embryonic development. To identify small molecules that induce CS in the embryonic neurons, we generated cerebral cortical neurons from GFP-expressing hESCs (H9, WA09) according to an established protocol (Qi et al., 2017; Figure S2). The ESC-derived cortical progenitors at Day 14 expressed SOX1 (86.7%) and OTX2 (87%), markers of cortical progenitors (Figure S2B). When differentiated to mature neurons in the presence of compound E that inhibits notch signaling and MEK inhibitor PD0325901 at Day 21, the majority of the cells expressed neuronal markers (MAP-2b 95%, TUBB3 95%; Figure S2C). Following treatment of the neuronal cultures with small molecules for 4 consecutive days, we assayed for CS hallmarks (Figure S2D). The criteria for positive molecules were defined by expression of CS markers without inducing obvious DNA damage and cell death. By using three different concentrations based on the half maximal inhibitory concentrations (IC50s) for each small molecule, we identified a concentration that did not cause cell death (Figure S2E). Romidepsin, O151, SBI-0206965, Lopinavir, Sodium Butyrate, SCR-7, and Phosphoramidon had a significant impact on the expression of all three readouts H3K9Me3 (mean ± SEM 1980 ± 22, 1957 ± 19, 1632 ± 15, 1806 ± 27, 1990 ± 18, 1908 ± 23, 2037 ± 24, respectively, compared to 2183 ± 14 in control), Lap2β (742 ± 6.4, 688 ± 6, 726 ± 5, 709 ± 8, 734 ± 5, 693 ± 7.7, 855 ± 7.5, respectively, compared to 789 ± 4 in control), and HP1γ (122 ± 3.6, 98 ± 0.5, 92 ± 0.3, 96 ± 0.64, 98 ± 0.5, 99 ± 0.5, 95 ± 0.5, respectively, compared to 108 ± 0.5 in control; Figure 3a,b; Table S2). Romidepsin induced a greater expression of HP1γ and Phosphoramidon induced greater Lap2β expression compared to the mean expression in the control group and were excluded from further experiments (Table S2). Among the remaining molecules, we found that neurons treated with actinomycin D, etoposide, temozolomide, and hydroxy-urea showed higher H2A.x expression compared to the control group (Figure 3c,d), suggesting that these molecules caused significant DNA damage, promoting us to exclude these molecules from further screenings. Five molecules (O151, SBI-0206965, Lopinavir, Sodium Butyrate, and SCR-7) were selected for further analysis.

Chemical induction of CS in hESC-derived cortical neurons. (a) Frequency distribution of high-content imaging data for H3K9Me3, Lap2β, and HP1γ proteins in cortical neurons. The dashed red line is control, and top seven molecules for each protein marker are shown in the graph. (b) Mean difference for signal intensity of all 25 small molecules depicted as mean ± 95% confidence intervals compared to the DMSO control. The zero line means no difference compared to the control and if the difference does not touch the reference line then changes in expression are significant. (c) Confocal images of phospho-Histone H2A.X (Serine 139) in the H9-GFP cortical neurons treated with Etoposide, Actinomycin D, and DMSO as control (Scale bar = 50 μm). (d) Quantification results for the number of positive foci for phospho-Histone H2A.X (Serine 139) per nucleus in cortical neurons treated with different small molecules. (n = 3, ns: not significant, *p < 0.05, **p < 0.01, ***p < 0.001 one-way ANOVA with Dunnett's multiple comparison test)
Our next step was to identify whether any combination of these five small molecules induces CS in neurons. We used the single molecule treatment with SBI-0206965 (autophagy inhibitor) as a reference since it had greater performance in modulating all three readouts during the initial screening. In this set of experiments, we used 50% of the concentration that we used for the first set of experiments for molecules used in pairs and 70% reduction in triple combination to minimize cell toxicity. Results showed that most of the combinations had greater or similar effect to SBI (Figure 4a). Two of the combinations, SLO (SBI-0206965, Lopinavir, O151) and SSO (SBI0206965, Sodium Butyrate, O151), had a greater mean difference in H3K9Me3 and Lap2β expression compared with both DMSO (Control) and SBI-0206965 treated cells (p < 0.01).

Combinatorial effect of small molecules on CS in cortical neurons. (a) Different combination of five most effective molecules (O151, SBI-0206965, Lopinavir, Sodium Butyrate, SCR-7) tested on cortical neurons and mean expression of H3K9Me3, Lap2β, and HP1γ in treatment groups compared to the DMSO control. (b) A graph showing the period of SLO treatment on the expression of Lap2β, HP1γ, and H3K9Me3 at Day 14 after maturation and (c–e) high-content imaging quantification of signal intensity for each marker after SLO treatment. (f) Relative frequency distribution of different bins of signal intensity for Lap2β, HP1γ, and H3K9Me3 in cortical neurons treated with different small molecules. (g) Representative images of Western blot for all three markers in cortical neurons treated at Day 21 of differentiation and (h) their normalized protein expression to tubulin expression. (i) Immunostaining images of H9-GFP cortical neurons treated with MG-132 (proteasome inhibitor), SLO (SBI-0206965, Lopinavir and O151), and SSO (SBI-0206965, Sodium Butyrate and O151) and stained for Lamp2A (Lysosome membrane associated protein) and Proteostat dye for detection of protein aggregation (Scale bar = 100 μm), and (j) quantification of positive area in neurons for Lamp2A and Proteostat. (i, j) Young and aged iNs added for comparison with ESC-derived cortical neurons. (n = 3, ns: not significant, *p < 0.05, **p < 0.01, ***p < 0.001 one-way ANOVA with Dunnett's multiple comparison test). For Western blot quantification, unpaired Student's t test was performed
To determine the minimum period of treatment needed to induce stable CS, differentiated cortical neurons at Day 7 were treated with the SLO small molecules for different periods of time (treated at Day 7, Day 9, Day 10, Day 12, and Day 13) and the cells were analyzed at Day 14. Expression of H3K9Me3, HP1γ, and Lap2β indicated that 2–4 days of continuous treatment with SLO molecules resulted in the maximum effect (Figure 4b–f). This experiment showed that expression of H3K9Me3 and Lap2β at 5- and 7-day post-treatment recovered slightly but not to the level of the DMSO condition. Reduction in HP1γ level was more persistent following SLO treatment and stayed at a lower level compared to the control cells even at 5- and 7-day post-treatment (Figure 4b–f). Downregulation of all three senescence-related proteins, H3K9Me3, HP1γ, and Lap2β, was confirmed by Western blot in the differentiated cortical neurons treated with SLO at Day 7 (Figures 4g,h and S7A-D).
In addition to the CS phenotypes analyzed above, neuronal senescence is often accompanied by intracellular protein aggregation. We hence examined the effect of the top two small molecule combinations on protein aggregation with MG-132-treated cells (a proteasome inhibitor) as a positive control. Proteostat™ staining revealed protein aggregates in cells treated with SSO or SLO comparable to MG-132 condition which was colocalized by Lamp2-positive autophagosomes (Tukey's multiple comparison MG-132 p < 0.0001, SLO p < 0.004, SSO p < 0.035; Figure 4i,j). As additional controls, Proteostat™ and Lamp2 staining revealed more prominent protein aggregation in aged iNs than in the young iNs (p < 0.003). Our results show that the CS phenotype in neurons induced by small molecules is associated with intracellular protein aggregation, similar to the phenomena preserved in aged iNs.
Mitochondrial defects are associated with senescence in the directly reprogrammed neurons (Kim et al., 2018). We found that SLO-treated neurons showed a higher ROS level than the control cells, revealed by MitoSoX staining (Figure S3A,C). It is, however, much lower than that in cells treated with FCCP, a potent uncoupler of mitochondrial oxidative phosphorylation and inducer of cell apoptosis. In parallel, JC-10 assay showed that the SLO-treated cortical neurons had a lower mitochondrial membrane potential than untreated controls, but again not as low as that in the FCCP treated cells (Figure S3B,D). Accompanying with the functional changes was morphological alterations in the mitochondria when neurons were treated with SLO, including fewer branches and smaller area for SLO-treated cells, though statistically insignificant (Figure S3E–G). Thus, SLO-induced CS is accompanied by functional alterations in mitochondria, including depolarization and over production of ROS.
2.3 SLO-treated neurons express CS-related transcripts and pathwaysTo define CS-related changes in SLO-treated neurons, we performed RNA-seq analysis on cortical neurons treated with or without SLO. Principal component analysis (PCA) based on overall gene expression showed high similarity (clustering) among independently cultured neurons treated with SLO or among those without SLO treatment (controls), but a high degree of separation between the SLO-treated and the control groups (Figure 5a). When comparing our RNAseq data with iNs from both young (<30 years, eight samples) and aged (>60 years, nine samples; Mertens et al., 2015), we found that our cortical neurons are similar to young iNs, whereas the SLO-treated neurons clustered with aged iNs (Figure 5a). We further compared our SLO-treated cells to the aged (>60 year, N = 205) and young (<30 year, N = 128) brain samples available in PsychENCODE (Wang et al., 2018). The SLO-treated samples (orange dots) are clustered with the PsychENCODE aged group (red dots), whereas the CTRL samples (blue dots) are clustered with the PsychENCODE young group (green dots). Note that we used PC2 and PC3 for showing clustering since the first PC (PC1) likely captures potential major confounding factors between our study and PsychENCODE (Figure 5b). Together, our results indicate that the SLO-treated neurons resemble those in the aged human brain and those directly converted from aged fibroblasts.

RNAseq analysis on SLO-treated cortical neurons. PCA plot for SLO and CTRL samples as well as induced neurons (iNs) converted from both (a) aged (nine samples) and young (eight samples) fibroblasts and (b) the aged and young PsychENCODE samples (the old group (>60 years, N = 205) and the young group (<30 years, N = 128)). Boxplots for human aging scores association between SLO neurons and brain samples for (c) upregulated genes and (d) downregulated genes in the aged brains. (e) Smear plot represents each gene with a dot, the gray dots (below cutoff line) are genes with no change relative to the contrast direction, red dots denote upregulated transcripts in the control neurons (with decreased expression in the SLO-treated neurons), and green dots denote downregulated transcripts in the control neurons (upregulated in SLO-treated neurons), respectively, at an adjusted p-value (FDR) significance threshold of 0.05. For FDR correction, we used Benjamini–Hochberg method. The light blue dots are transcripts with FDR < 0.05 but have log expression change of less than 0.6. The x-axis (log2 fold change) is the effect size, indicating how much expression has changed with SLO treatment. (f) Heatmap clustering for 50 of the most differentially expressed genes with a p-value < 0.05 and a log (2) fold-change greater or less than 2. The Z-score of a given expression value is the number of standard deviations away from the mean of all the expression values for that gene. (g) All DEGs with a FDR < 0.05 and 0.6 ≤ logFC ≤ −0.6 are selected and tested for over- or under-representation of pathways in the gene list. Any significantly enriched WikiPathway pathways are ordered from most to least significant based on the p-value
We then looked at the human aging scores (−log10(p-value) for the genes that are associated with aging, see RNAseq in methods) of cortical neurons treated with SLO. We found that the upregulated genes in SLO-treated cells have higher human aging scores in the PsychENCODE aged group than the downregulated genes (Figure 5c, t test p < 2.2e−16). Similarly, the downregulated genes have significantly higher human aging scores in the PsychENCODE young group than the aged genes (Figure 5d, t test p < 2.2e−16). These results suggest that our SLO-treated neurons have a similar gene expression dynamic to that in the aged human brain in PsychENCODE.
Comparison between SLO-treated neurons and DMSO control neurons resulted in 271 differentially expressed genes (DEGs; FDR < 0.05, 0.6 ≤ logFC ≤ −0.6) with 190 genes downregulated and 81 genes upregulated upon SLO treatment (Figure 5e; Table S3). These DEGs are also present in the gene list that are significantly modulated by age in PsychENCODE (Table S4). In our SLO DEG list, GABA receptors are among the most downregulated genes whereas histone variants are upregulated (Figure 5f). Pathway analysis for DEGs in the SLO-treated neurons revealed that neurotransmitter receptor signaling and GPCR signaling are downregulated whereas pathways in the histone modification (especially histone variants) are upregulated (Figure 5g).
Premature aging syndromes that are associated with mutations in LMNA or WRN genes resemble normal aging in terms of gene expression (Dreesen & Stewart, 2011; Kyng et al., 2003). Overexpression of mutant Lamin A/C (Progerin) in normal neurons causes aging phenotypes (Miller et al., 2013). Interestingly, the SLO-treated neurons exhibited an upregulated pathway (WP4320) that shares 11 genes (30% of total genes in the pathway) involved in Hutchinson–Gilford Progeria syndrome (Figure 5g). They included histone variants, several of which are involved in the histone modification pathway (WP2369). Other transcripts that are upregulated in SLO-treated cells included insulin receptor substrate 1 and 2 (IRS1 and IRS2), pro-apoptotic genes (FOXO3, BAD, and BCL2L11), nutrients sensing transcripts (EIF4EBP1, TSC2, and EEF2) and downstream kinase molecules (PIK3R2, ELK1, PTPRF, MAPK7, AKT1, MAP2K2, PLCG1, CRTC1, and JUN), and other transcripts (SHC2, RAB3A, DOCK3, RELA, NCK2, RACK1, SH2B1, LINGO1, STAT5B, EGR1, and SQSTM1). Other downregulated transcripts in SLO-treated cells included AMPA and NMDA receptors (GRIA1, GRIA2, GRIA3, and GRIN2B), both trkB and trkC receptors (NTRK2 and NTRK3) and their downstream calcium signaling molecules (NFATC4, CAMK2A, and CAMK4), MAPK responsive transcripts (MAP2K1, KIDINS220, PRKAA2, and PPP2CA), and other transcripts (GABRB3, MEF2C, SHC3, RASGRF1, PIK3R1, CDC42, CDH2, CNR1, SPP1, EIF4E, NSF, PTPN11, DLG1, and APC). The transcriptome data suggest that the SLO-treated neurons resemble those from aged human cortex and premature aging samples.
2.4 Induction of CS accelerates disease phenotype manifestation in ALS MNsNeurodegenerative diseases such as amyotrophic lateral sclerosis (ALS) usually manifest symptoms after the 5th decade of life. We hypothesized that induction of CS in ALS iPSC-derived neurons would accelerate the presentation of disease phenotypes. We used the TARDBP mutant (298S) iPSC line generated from an ALS patient and its isogenic cell line (298G) produced by correcting the mutation using CRISPR (Figure S4). Both mutant and corrected iPSCs were differentiated to spinal MNs using our previously published protocol (Chen et al., 2014; Du et al., 2015; Figure 6a), and the MNs were treated with the SLO cocktail for 4 days. As expected, addition of the small molecules at Day 25 and assayed at Day 28 did not significantly alter the percentage of cells expressing cleaved caspase 3 (Figure 6b), indicating no obvious cytotoxicity for SLO and SSO treatments (control = 26.9 ± 1.3, SLO = 25 ± 2.4, SSO = 24.7 ± 4.9) whereas we found significant number of caspase 3 positive cells in MG132 treated neurons (MG132 = 36.92 ± 1.33).

Phenotype presentation by SLO-treated motor neurons derived from TARDBP mutant iPSCs. (a) Differentiation protocol used for generating MNs from TDP-43 298S mutant and 298G isogenic iPSCs. (b) Immunostaining and quantification for cleaved caspase 3 and alpha internexin proteins in cultured MNs treated with SLO, SSO and MG-132, at Day 28. (c) High-content imaging for H3K9M3 and Lap2β in both TDP43 298G isogenic control and 298S mutant following SLO treatment (Mean of SLO treatment compared to the control group with DMSO). (d) Representative phase contrast image of MN cultures from both control and mutant ALS neurons treated with SLO. (e) Immunostaining images for alpha-internexin, Proteostat, phosphorylated neurofilament heavy proteins in control and mutant MNs treated with SLO; right panel shows higher magnification images of control and mutant MNs treated with SLO (Scale bar = 200 μm, for higher magnification images scale bar = 50 μm). (f) Quantification result for phosphorylated neurofilament aggregates and (g) Proteostat-positive protein aggregations. (h) Quantification of phosphorylated TDP43 protein in the nuclei area across all groups and also (i) quantified nuclear TDP43 protein expression for both 298G, S neurons and SLO-treated neurons. (j) Mitochondrial membrane potential (JC10 assay) of ALS-iPSC-derived MNs treated with SLO compared to the healthy isogenic control cells and isogenic cells treated with FCCP. (n = 3, ns: not significant, *p < 0.05, **p < 0.01, ***p < 0.001 one-way ANOVA with Dunnett's multiple comparison test, for JC10 assay data were collected using 15,000 cells per group from three independent experiments. Statistical analysis was performed using one-way ANOVA, Tukey post hoc test (****p < 0.001))
We then examined if the MNs treated with SLO display CS-like phenotypes as we observed in fibroblasts and cortical neurons. Both 298G and 298S MNs showed a reduction in the expression of H3K9Me3 and Lap2β following SLO treatment (Figures 6c and S6A), indicating that SLO treatment induces CS. Both cell lines responded at the same level to the SLO chemicals, and difference in H3K9Me3 and lap2β signal intensity was not significant (Figure 6c). We then asked if induction of CS accelerates neuronal degeneration. By Day 32, the 298S MNs treated with SLO showed axonal swellings, a sign of axonal degeneration whereas 298G MNs showed relatively intact neurites (Figure 6d). Proteostat staining was increased in SLO-treated cells, especially in the 298S MNs. ALS MNs, when treated with SLO, were positive for phosphor-TDP43 and Proteostat (Figures 6e and S5). Immunofluorescence for phosphorylated neurofilament, a marker for axonal degeneration and turnover, was also significantly increased in the SLO-treated 298S than in non-SLO-treated 298S and SLO-treated 298G MNs. Under higher magnification, Proteostat-positive aggregates were observed along the axons and were positive for both α-internexin and phosphorylated neurofilament (Figures 6e and S5). Quantification of the aggregates showed a significant increase in p-NFH aggregates (1.22 ± 0.31 in 298G compare to 4.26 ± 0.92 in 298S) and Proteostat-positive aggregates (4.49 ± 0.38 in G298G compare to 7.86 ± 1.06 in G298S) in 298S ALS mutant MNs than the 298G isogenic control MNs that were treated with SLO (Figure 6f,g). Following SLO treatment, ALS MNs had significantly more p-TDP43 signal compared to the isogenic control and cells that are not treated with SLO (Figures 6h and S5). We also detected TDP43 signal permeation from nucleus to the cytoplasm area in the ALS MNs and neurons treated with the SLO (Figures 6i and S5). Interestingly, neurite swelling contained p-TDP43 proteins that are colabeled with Proteostat and other neurofilaments are more pronounced in the ALS MNs treated with the SLO (Figures 6e and S5).
One of the hallmarks of neurodegeneration is mitochondrial deficit. Assay with JC-10 staining showed that ALS-MNs had a lower mitochondrial membrane potential than the isogenic control MNs. Treatment with SLO lowered the membrane potential for control neurons but not further for the ALS-MN (Figure 6j). Morphological analysis with Mitotracker staining showed that isogenic control MNs treated with SLO had shorter and smaller mitochondria than those without SLO treatment, but SLO treatment had no further effect on ALS MNs (Figure S6B-D). Given that mitochondrial DNA is generally not reprogrammed during iPSC generation, these results suggest that the mitochondrial phenotypes are present in ALS-iPSC-derived MNs and that SLO treatment does not appear to exacerbate the mitochondrial phenotypes beyond what are present in ALS-MNs.
2.5 Autophagy Induction clears up protein aggregation and improves neurite healthThe fast and consistent presentation of disease-relevant phenotypes in SLO-induced cultures makes them amenable for testing therapeutic agents. We examined the effects of molecules on protein aggregation and neurite swelling in the SLO-treated ALS MN cultures, including the current ALS medication Edaravone, autophagy activators STF-62247, SMER28, Flubendazole, and the peptide Tat-Beclin, and KU-60019, a molecule identified from our initial cell toxicity screening as neuroprotective. In addition, amiodarone was used as an unrelated hit. MNs from both ALS (298S) and isogenic (298G) iPSCs at Day 28 post-differentiation were treated with SLO and then the compounds were added separately 24 h later, and cells were analyzed at Day 32 (Figure 7a). SMER28 and Tat-Beclin decreased proteostat-positive aggregates in both ALS (37% ± 14 and 62.5% ± 5.5) and isogenic MNs (38% ± 11.7, 66.9% ± 5 for 298G) as compared to SLO-treated control. Edaravone and KU-60019 reduced the level of Proteostat, more so on ALS cells (to 26% ± 6 and 17% ± 0.7) than the isogenic cells (64% ± 10.4, 30% ± 13.6 for 298G). MNs treated with STF-62247 and Flubendazole showed more aggregates in 298G cells compared to the SLO-treated cells (179% ± 29, 143 ± 26.8) and no improvement in 298S cells. Amiodarone did not improve protein aggregation (Figure 7b).

Testing molecules that rescue the disease phenotype in ALS MNs. (a) MNs from TDP-43 G298S mutant and G298G isogenic iPSCs treated with SLO to induce CS phenotypes and 24hr later candidate molecules SMER28, EDARAVONE, and KU-60019 were added to the culture and cells stained with Proteostat dye for protein aggregation and alphaInternexin for visualizing neurites. (b) Quantification of immunostaining images positive for Proteostat and (c) neurite swellings. (n = 3, Scale bar=100 μm)
Neurite swelling is one of the obvious morphological changes in ALS MNs following SLO treatment. SMER28 and EDARAVONE significantly reduced the number of swellings to the level of isogenic control cells (Figure 7c). However, KU-60019 treatment did not improve the swelling phenotype to the normal level (Figure 7c) despite significant reduction in protein aggregation (Figure 7b). Other molecules did not show significant improvement in 298S MNs or even induced more swellings in the 298G control cells (Figure 7c). Thus, EDARAVONE and SMER28 can reduce both protein aggregation and neurite swellings in 298S TDP43 mutant cells and were beneficial for MN health in our CS culture system.
3 DISCUSSIONMost neurodegenerative diseases are concurrent with aging (Baker & Petersen, 2018; Bickford et al., 2017; Duncan, 2011; Sawada et al., 2008). Hence, recapitulating CS in stem cell-derived neurons could expand the capacity of the iPSC model to study disease mechanisms (Mertens et al., 2018; Vernadakis & Fleischer-Lambropoulos, 2000). By using H3K9Me3, HP1γ, and Lap2β as readouts and screening for chemicals/pathways that induce CS in neonatal fibroblasts and iPSC-derived cortical neurons, we developed cocktails of small molecules that induce CS in the cortical neurons. This CIS approach was validated in MNs derived from ALS patient iPSCs. Importantly, CIS enhanced the presentation of disease-related phenotypes. This CIS strategy will likely enable more effective iPSC-based modeling of age-related degenerative diseases and enable better therapeutic target design.
Cellular senescence across different cell types shares features including mitochondrial dysfunction, DNA damage, P16 expression changes, and epigenetic marks for gene silencing (Kim et al., 2018; Madabhushi et al., 2014; Rubinsztein et al., 2011; Satoh et al., 2017). These alterations ultimately result in age-related changes at the cellular level, including changes in cell size, shape and metabolism, proliferation arrest, and telomere erosion (Hernandez-Segura et al., 2018; Lopez-Otin et al., 2013; Petrova et al., 2016). In mitotic cells like fibroblasts, expression of P16 accompanies proliferation arrest and induces senescence (Coppe et al., 2011; Rayess et al., 2012). P16 activation by Palbociclib in our study is one of the most efficient pathways in CS by blocking CDK4/6 and proliferation of fibroblasts, causing senescence. Other pathways in our study with fibroblasts are related to the DNA repair, DNA synthesis, and DNA alkylation pathways; all related to cell division and telomere attrition. Surprisingly, none of the sirtuin inhibitors induced senescence in fibroblasts or neurons despite their effects on aging (Bonda et al., 2011; Satoh et al., 2017). This may reflect differences between cell types or insufficient treatment with the inhibitors.
In post-mitotic cells like neurons, protein quality control, including proteasome and autophagy processing, is more important in CS progression (Pan et al., 2008; Scotter et al., 2014; Zhang et al., 2017). This is reflected in our study showing the powerful CS-inducing effect of autophagy inhibitors (SBI-0206965). Faulty autophagosomes could not clear impaired mitochondria and unfolded protein debris, leading to lack of mitochondrial turnover and producing more oxidative stress (He et al., 2013; Wyss-Coray, 2016). Oxidative stress generates ROS and accounts for higher DNA mutations, which is ultimately related to CS (Campos et al., 2014; Lo Sardo et al., 2017). Similarly, we found that inhibition of DNA glycosylase (OGG1), important in detecting and removing oxidized nucleotides in genomic DNA, exacerbates CS phenotype in neurons but not in fibroblasts. Two of the three small molecules in SLO, DNA glycosylase inhibitor (O151) and HIV protease inhibitor (Lopinavir), modulate CS phenotypes in neurons, indicating that base excision repair (BER) pathway is critical for neuronal health and is linked to neurodegenerative diseases (Leandro et al., 2015; Maynard et al., 2015). Lopinavir also inhibits ZMPSTE24 (Coffinier et al., 2007; Mehmood et al., 2016), thereby blocking lamin A biogenesis and leading to an accumulation of prelamin A. ZMPSTE24 deficiency in humans causes an accumulation of prelamin A and leads to lipodystrophy and premature aging (Afonso et al., 2016; Spear et al., 2018; Wang et al., 2016), which perhaps causes senescence phenotype in our cultured neurons. We used three different endopeptidase inhibitors Phosphoramidone (a general metalloendopeptidase), Lopinavir (zinc metalloprotease inhibitor), and GGTI-298 (a geranylgeranyltransferase I (GGTase I) inhibitor), and only Lopinavir induced senescence in cortical neurons in all three markers. Interestingly, none of the endopeptidase inhibitors induced senescence phenotype in fibroblasts, indicating that neurons are more sensitive to the activity of endopeptidase, perhaps for the processing of other lamin proteins rather than just for Lamin A (Jung et al., 2012; Yang et al., 2015).
Information on CS derives largely from studies on mitotic cells. The gene expression pattern of our SLO-treated neurons, revealed by transcriptomic analysis, resembles that of iNs and aged brain. In particular, our in-vitro neuronal senescence system, despite the lack of many other cell types that are normally present in the human brain, resembles the aging cortex samples as indicated by the substantial overlap of age-related transcripts between our CIS neurons and aged human brain tissues (Wang et al., 2018). These transcription profiles may be more specific to CS in neurons. For example, transcripts that are involved in neurexin/neuroligin complexes at synaptic membrane assembly and neurotransmitter release from GABA, glutamate, and cholinergic systems are common between aged brains and SLO-induced senescence. Neurexin expression declines with age and causes decrease in synaptic density and cognitive decline (Konar et al., 2016; Kumar & Thakur, 2015). Other transcripts like CREB regulated transcription coactivator 1 (CRTC1) transcription coactivator of CREB1 (Paramanik & Thakur, 2013), which show significant change in our SLO-treated cortical neurons, also contribute to brain aging and neuronal senescence. Some of other molecules such as p62 (SQSTM1) have multiple function and contribute to neurodegeneration by binding to the ubiquitin molecules that are marked for degradation and by binding to autophagy molecule LC3-II (Ma et al., 2019). In addition, our CIS neurons share several histone variants with the progerin effect in the progeria syndrome. Histone variants are one of the most affected transcripts during CIS in the cortical neurons. Histone variant exchange, by regulating expression of age-related genes (Gevry et al., 2007) and/or chromatin organization (Flex et al., 2019), is one of the mechanisms behind CS and aging. Thus, our CIS recapitulates aspects of premature aging effects primarily at the epigenetic level.
A major driving force behind the development of CIS is to enable effective and reliable modeling of age-related diseases using human stem cells. We and others have shown that some aspects of neurodegenerative changes such as ALS may be recapitulated by strictly controlling the neuronal differentiation process, prolonged maturation, and undergoing stress (including culturing under a basal condition without trophic factors and medium changes; Chen et al., 2014; Qian et al., 2017; Sances et al., 2016). Such manipulations over a long term add variables to the system, making stem cell-based disease modeling more difficult. MNs from patients with TARDBP mutations have increased levels of soluble and detergent-resistant TDP-43 and show decreased cell survival, suggesting that this model is representative for ALS pathology (Bilican et al., 2012; Fujimori et al., 2018). However, neither increase in insoluble TDP43 nor its mislocalization phenotype is repeated in a recent study (Klim et al., 2019). Similarly, dopamine neurons from PD iPSCs exhibited mitochondrial dysfunction and oxidative stress, changes in neurite growth and morphology, synaptic connectivity, and lysosomal dysfunction (Kouroupi et al., 2017; Monzio Compagnoni et al., 2018; Reinhardt et al., 2013; Sanchez-Danes et al., 2012), but hallmark pathology like protein aggregation and Lewy body formation are rarely observed (Kouroupi et al., 2017; Mishima et al., 2018; Monzio Compagnoni et al., 2018; Reinhardt et al., 2013). These inconsistencies may be due to the different protocols used and the long-term cultures that are necessary to mature the stem cell-derived neurons. The current CIS approach enables an early and consistent presentation of disease-relevant phenotypes, including protein aggregation and axonal degeneration in TDP43 mutant MNs. Since the cocktails induce CS in different neuronal types, it is likely that the CIS approach may promote phenotypic presentation in other age-related diseases using iPSCs.
Since our CIS method enables faster and consistent presentation of disease phenotypes from iPSC-derived neurons, it is also useful for establishment of drug testing platforms. As a proof of principle test, we found that the current ALS medication Edaravone and one of the many autophagy activators SMER28 but not others mitigate protein aggregation and neurite swelling in ALS iPSC-derived MNs, highlighting the utility of the system.
3.1 Limitations of the studyOur CIS method induces CS in a short period (after 4 days of treatment) without a need of genetic manipulation. However, it may be desirable to induce senescence
Comments (0)