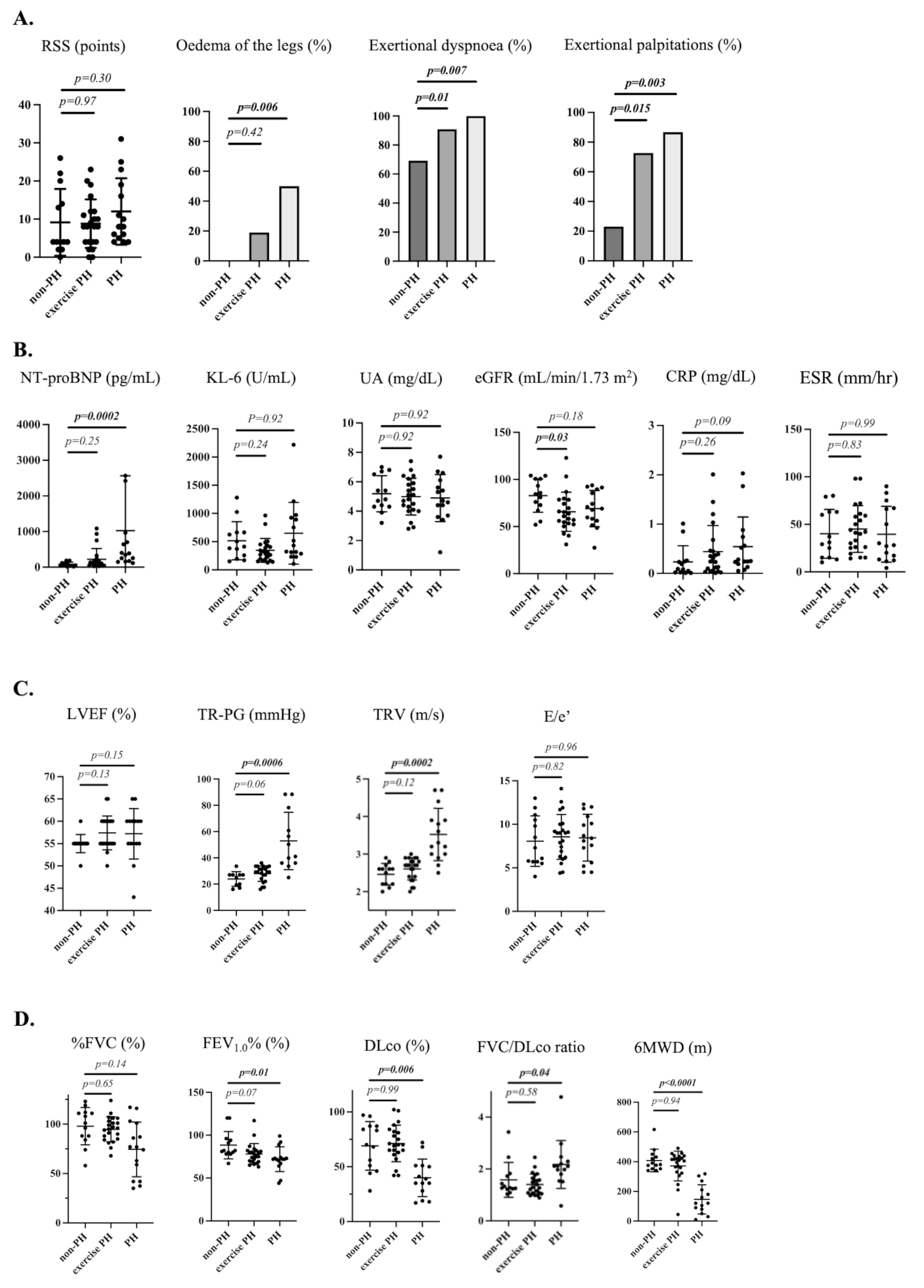
Cadherins can play a crucial role in fibroblast function by regulating cell-cell adhesion and maintaining tissue architecture. They can serve as signaling proteins and are involved in cellular junctions, migration, and maintaining tissue homeostasis [8, 9, 29]. In RA, CDH11, in particular, has been implicated as a key protein required for FLS homotypic aggregation in the synovial intimal lining [7, 8]. To understand if there are broader functions of the cadherin family in synovial biology, we evaluated the expression of other cadherins in FLS. Unexpectedly, we identified another cadherin, namely CDH6, as a gene of interest in RA. Notably, CDH6 is expressed at higher levels in RA FLS compared with OA FLS and promotes an aggressive phenotype, potentially through a mechanism involving migration and cell survival. These studies suggest that targeting CDH6 in RA might have potential clinical benefit by attenuating the aggressive behaviors of FLS. This molecule does not appear to play a major role in immune responses, possibly enabling its use in combination with agents that affect host defense. In addition, this approach could mitigate fibroblast-mediated structural damage when it progresses despite suppressed inflammation or could be used as a biomarker to identify patients with fibroblast activation.
CDH6 is a type II cadherin and is highly expressed in kidneys, brain, and liver. It is overexpressed in a variety of tumors, including renal carcinomas, gliomas, thyroid cancer, gastric cancer, and breast cancer [5, 10,11,12, 30, 31]. Its expression also correlates with lymph node invasion and metastasis, serving as a marker for a poor prognosis [32]. Our study shows for the first time that CDH6 expression is also elevated in RA FLS, which display many features of aggressive tumor cells, such as enhanced invasiveness and survival [5, 26, 31, 33]. In synovial tissue, its expression is observed mainly in FLS, but it is also displayed by MLS in intimal lining regions, as well as other cell types in sublining regions. The interaction between FLS and MLS through adhesion molecules was seen previously, such as the VCAM-1 and integrin α4β1, which are known to contribute to the structure of the synovium in vivo [7]. Interestingly, it was observed that CDH6 is associated with immune infiltration and cytokine expression in glioma, suggesting that the CDH6 presence may facilitate the interaction between FLS and MLS, potentially contributing to the hyperplastic lining and chronic inflammation.
CDH6 has two splice variants: a long form that includes an extracellular domain with a cytoplasmic domain and a short form lacking this domain [26]. Of interest, certain cadherins have a signaling role by interaction with downstream molecules through their cytoplasmic domains, such as E-CDH and N-CDH [27, 28, 34]. In thyroid tumor cells, both CDH6 isoforms are expressed [26, 31]. However, in FLS the long form is much more abundant, suggesting that intracellular interactions of CDH6 might contribute to the function of CDH6. Interestingly, we also found abundant CDH6 intracellular distribution in FLS, including localization to the nucleus and perinuclear regions, further suggesting CDH6 might contribute to intracellular signaling mechanisms. The intracellular distribution could also explain why the CDH6 antibody did not block cell growth compared with siRNA knockdown in FLS in our studies. In contrast, the CDH6 antibody did inhibit migration, indicating that both the extra- and intra-cellular domains of CDH6 play roles in FLS biology.
Increased expression of CDH6 in RA FLS is associated with specific epigenetic marks associated with other pathologies. For example, a high level of tri-methylation on lysine 27 of histone H3 (H3K27me3) at the CDH6 gene locus is observed in multiple myeloma cells, and this modification correlates with low CDH6 expression [35]. To help understand increased expression in RA, we reviewed epigenetic landscape data and discovered that chromatin accessibility and histone modifications, particularly H3K27ac, are enhanced in RA FLS compared with OA FLS. H3K27Ac is closely associated with open chromatin and gene expression, which likely explains the higher CDH6 transcription in RA FLS. To confirm this, we treated FLS with a histone deacetylase inhibitor, which equalized CDH6 expression between RA and OA FLS. These data suggest that histone acetylation plays a critical role in the differential regulation of CDH6 in RA.
In addition to epigenetic markers, we also showed that CDH6 is induced by TGFβ in RA FLS. Previous studies have shown that CDH6 expression is increased by TGFβ during epithelial-to-mesenchymal transition in cancer, promoting invasiveness in tumor cells [10, 26]. TGFβ is a multifunctional cytokine involved in diverse biological responses, such as angiogenesis, cell proliferation, differentiation, migration, and apoptosis, and it plays a pivotal role in fibrosis and pathogenesis associated with RA [36, 37]. Our data suggests that CDH6 may act as a downstream mediator of TGFβ signaling in the RA synovium. However, further investigation is necessary to understand this mechanism.
CDH6 function has been implicated in cancer cell migration, invasion, and cell growth [5, 13, 33, 38]. For example, CDH6 knockdown decreases migration and invasion in ovarian and renal cancer cell lines, which was attributed to reduced AKT and ERK activation [31]. We showed that CDH6 regulates similar functions in RA FLS that might contribute to their aggressive behaviors. However, in RA FLS, mechanism studies did not implicate AKT, ERK, or cell cycle abnormalities. Surprisingly, autophagy in FLS was also not regulated by CDH6, even though that process has been implicated in other cell types [11]. However, we did demonstrate that CDH6 knockdown modestly decreases cell survival, offering an explanation for its effect on cell growth.
These studies have some limitations. First, our findings are based on in vitro experiments using primary FLS cultures, which might not reflect the situation in situ. Even though all FLS lines express CDH6, there is variability in CDH6 expression among FLS lines. Therefore, we used multiple lines to ensure that we had representative samples. Also, we were able to determine the effect of CDH6 on cell survival, but the precise molecular mechanism remains unknown. We suspect that it is related to intracellular functions of the long form. The role of CDH6 in joint development or structure is also an area that is not fully explored. Our studies focused on in vitro assays, and the effect of CDH6-directed approaches in pre-clinical arthritis models would be helpful. However, the interpretation of these experiments might be difficult because mouse CDH6 has distinct functions compared with the human protein. For example, CDH6 interacts with αIIb/ß3 on platelets and is associated with thrombosis in humans but operates through a different non-platelet mechanism in mice [39]. Finally, our studies focused on patients with longstanding disease requiring arthroplasty, although it would be interesting to evaluate early-stage disease to determine if outcomes correlate with CDH6 expression and function.
In conclusion, our findings reveal that CDH6 is differentially expressed in RA FLS over OA FLS and plays a significant role in RA FLS pathogenic behavior, sharing regulatory and functional similarities to its role in cancer. The increased differential RA FLS expression may contribute to FLS survival and migration in situ, contributing to RA pathology and chronicity. Disrupting CDH6-mediated cell interaction may reduce synovial hyperplasia in RA. CDH6 has been shown to be an attractive target for treating certain cancers. For example, a phase I study of HKT288, a CDH6-targeting antibody-cytotoxic drug conjugate, was terminated due to neurologic adverse events [15]. Another phase 1a dose-escalation has been reported with another antibody-payload approach [16]. Given CDH6 involvement in aggressive phenotypes in RA, these CDH6-targeted approaches might be useful in RA as well.



Comments (0)