
Remember me
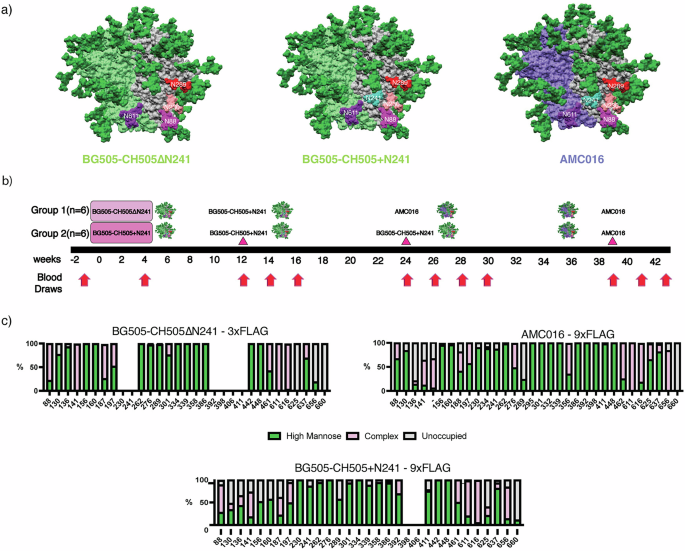

Previous FP studies with non-glycosylated trimers around the FP have resulted in Ab responses biased towards the glycan deletions13,23. Instead, we hypothesized that a more conservative glycan deletion approach would induce FP-specific responses capable of broad HIV-1 coverage. In this study, we primed rhesus macaques (RMs) using chimeric trimers based on BG505 and CH505 with the V1, V2, V3, and V5 loops of CH50527, a clade C virus, grafted onto a clade A BG505 trimer background as wild-type BG505 does not carry two of the four canonical FP glycans at positions N241 and N23028,29. BG505 also lacks a conserved glycan at position 289, which opens up an epitope previously shown to be immunogenic, so we introduced a glycan at position 289 to mitigate the immunodominance of this glycan hole epitope (Fig. 1a)29. In the experimental group, the chimeric priming immunogen presented three of the four FP glycans: N611, N88 and N230 but lacking in the N241 glycan (BG505-CH505ΔN241) (Fig. 1a). Alternatively, the control group was primed with an autologous, chimeric immunogen with all four canonical FP glycans present, including N241 (BG505-CH505 + N241). For the priming period, both groups of animals were given their respective soluble antigens and adjuvants via continuous delivery via an osmotic pump over a four-week period at a dose of 100 ug of antigen and 750 ug of SMNP adjuvant (Fig. 1b)23,24,30. All subsequent immunizations were given as bolus with the same dosages. The animals were first boosted with the BG505-CH505 + N241 immunogen at week 12 and later boosted with a heterologous, clade B trimer, AMC01631, but with BG505 FP sequence grafted, to further immunofocus on the FP and suppress boosting of pre-existing Abs against off-target epitopes (Fig. 1b). The AMC016 trimer is highly glycosylated and therefore suitable to suppress BG505-CH505-specific off-target and glycan hole-directed pAb responses29,32,33. AMC016 has previously been shown to induce strain-specific, neutralizing Abs in rabbits after autologous immunizations31.
Fig. 1: FP targeting Immunogens and Immunizations.
a Surface representations for BG505-CH505ΔN241 and BG505-CH505 + N241, which are BG505 (Clade A)/CH505 (Clade C) chimeric trimers. The AMC016 trimer is a Clade B isolate. FP glycans are depicted as follows: N88 glycan in magenta, N230 glycan in salmon, N241 glycan in turquoise, and N611 glycan in purple. The N289 glycan is depicted in red. b Immunization schedule. Osmotic pump immunogen delivery period is shaded in pink. Bolus immunizations are depicted with triangle indicator. c Glycan Analysis of immunogens used in this study and impacts of Flag tag length on glycan occupancy.
To shield an otherwise immunodominant epitope at the base of soluble Env trimers, non-native N-glycans were introduced at positions 656 and 660 (Fig. 1c) in all the constructs. Previous research has shown that membrane-bound trimers have higher glycan occupancy and improved glycan processing than soluble trimers due to longer retention in the ER/Golgi34,35. Here we employed an analogous strategy to extend the C-terminus of Env by introducing a Furin cleavage site (RRRRRR)36 after residue 664 of gp41, followed by a GS-rich linker to a 3x-Flag-Tag (priming immunogens) or a 9x-Flag-Tag (boosting immunogens). This tag increases the translation time and is subsequently cleaved off by Furin protease and is therefore not present in the final immunogen (Fig. S1a).
When a 3x-Flag-Tag was used, the introduced glycan at the C-terminus of gp41, N656 was less than 40% occupied while N660 remained completely unoccupied (Fig. 1c). When a 9x-Flag-Tag was used, the glycan at N656 was ~75% occupied, while glycan occupancy at N660 remained 0%. In these constructs there was also a large change in the glycan occupancy at canonical gp41 glycan at 625, which historically has low occupancy when expressed as a soluble trimer truncated at position 664 with a stop codon37. In the 3x-Flag-Tag constructs, the glycan at position 625 was completely unoccupied. However, in the autologous and heterologous boosting immunogens, which have 9x-Flag-Tags, there was a significant increase in glycan occupancy at position 625 to 40% in the BG505-CH505 + N241 autologous boost and 98% in the AMC016 heterologous boost (Fig. 1c). Although a PNGS sequon at N230 was present in the Env sequence for the BG505-CH505ΔN241 construct, mass spec analysis of this glycan site was unable to determine the occupancy levels of this FP glycan and so the occupancy of this site remains unknown.
Historically, BG505 SOSIP.664 has introduced cysteine residues at positions 501 of gp120 and 605 of gp41, forming an intra-protomer disulfide bond that stabilizes recombinant native-like Env trimers28. Despite this Env stabilization effort, antibody-mediated immunogen disassembly has been observed after repetitive autologous trimer immunizations38. Therefore, we introduced a new, inter-protomer disulfide linkage in the BG505-CH505ΔN241 and BG505-CH505 + N241 constructs, 501C-L663C, to prevent in-vivo trimer disassembly. Notably, this disulfide prevents binding of RM20A3, a base binding mAb isolated from macaques that were repeatedly immunized with BG505-based constructs (Figs. S1b and c)12,38,39. The AMC016 construct has the cysteines at positions 501 of gp120 and 664 of gp41, which also prevent RM20A3 binding (Figs. S1b and c).
NsEMPEM detected FP response after heterologous boostingPrior studies using EMPEM to map pAbs against different epitopes of the HIV Env protein provide a basis to evaluate the success of our immunofocusing approach12,26,40. Here, we used nsEMPEM to analyze pAb responses at post immunization timepoints using probe antigens matched to the most recent immunogen (Fig. 2a and Fig. S2a)25. At week 4 only base binders were bound to the probing antigens, in this case the priming immunogens, in each group (Fig. 2a and Fig. S2a). This result aligns with previous observations showing base responses are consistently elicited early in soluble Env trimer immunizations in animal and human studies12,23,24,25,26,40,41.
Fig. 2: Humoral response analysis via ELISA and nsEMPEM.
a nsEMPEM dot plot results after each immunization. b EMPEM figure legend from multiple views. c FP-ELISA binding for time points also analyzed via nsEMPEM – experimental group (dark grey, circle indicators) versus control group (light grey, triangle indicators). Animals RUu18, RQk18, and LJ66 are highlighted in green, pink and orange, respectively. d Autologous trimer ELISA showing serum recognition of BG505-CH505 + N241 trimer from control group (red) as well as experimental group (blue). (X) mark indicates immunization. e Heterologous trimer ELISA showing serum recognition of Heterologous AMC016 trimer from control group (red) as well as experimental group (blue).
After the initial priming immunizations, both groups were boosted with an autologous, chimeric immunogen with all four FP glycans present, BG505-CH505 + N241, the same immunogen given to the control group for priming (Fig. 1). Week 14 nsEMPEM analysis revealed more epitopes detected relative to week 4 (Fig. 2a and Fig. S2a). In addition to base binding pAbs, we observed pAbs bound to the C3/V5 region and V1/V2 region in most animals in both groups. One animal in the experimental group, LJ66, also developed a gp120/gp120 interface (IF) response (Fig. S2a). One animal in the control group, RUu18, revealed an FP response (Fig. S2a), however, follow-up cryoEM (discussed below) and FP-ELISA (Fig. 2b, green triangle) demonstrated that the FP response observed in RUu18 via nsEMPEM did not specifically target the FP.
While FP pAbs were detected as early as six weeks in our earlier 2018 NHP study23, FP targeting pAbs were not observed via nsEMEPM until 4 weeks after the heterologous AMC016 trimer boosts at weeks 24 and 39 in the majority of animals within the experimental and control groups, respectively, in this study (Fig. 2a and Fig. S2a). This suggests removal of only the N241 glycan around the epitope does not induce a robust priming immune response against the FP. Although FP responses were not observed at the earlier time points, FP-ELISA data suggested FP-recognizing Abs were elicited at low levels during chimeric BG505-CH505 construct immunizations (Fig. 2c and Fig. S2b).
Week 28 nsEMPEM analysis of the experimental group animals revealed base and FP responses, and one animal, LJ66, additionally showed a gp120/gp120 IF response (Fig. 2a and Fig. S2a). The control group, which had received three immunizations of the BG505-CH505 + N241 antigen by this point, still exhibited base, V1/V2/V3, and V5 responses in all the animals. Two animals in the control groups revealed gp120/gp120 IF responses and FP responses, respectively, via nsEMPEM analysis. While our data suggests that a single canonical FP glycan deficiency on Env does not prime a robust FP response, the results from the heterologous AMC016 boost were encouraging.
To distinguish if post-AMC016 boost FP elicitation in the experimental group was due to epitope accessibility in the priming immunogens or introduction of the heterologous boosting immunogen, both groups were immunized a fourth and final time with the heterologous AMC016 trimer at week 39. NsEMEPM analysis of week 43 plasma samples in the control groups showed elicitation of FP-targeting Abs in five of the six control animals (Fig. 2a). The experimental group, which had received two immunizations of the AMC016 trimer, elicited FP and base responses, as observed in week 28, however most of the animals now also exhibited AMC016-specific C3/V5 responses. Strain-specific responses after autologous immunizations but not after heterologous immunizations is a commonly observed phenomenon in multiple viral vaccination platforms42,43. Although nsEMPEM suggested both groups showed FP-directed responses after heterologous boosting, FP-ELISA showed that the experimental group had significantly higher recognition of the FP, suggesting that the N241 glycan hole in the BG505-CH505ΔN241 immunogen did impact FP-specific responses later boosted by the heterologous AMC016 trimer (Fig. 2c and Fig. S2b), however, differences in number of autologous and heterologous boost administrations and timing of heterologous boosting post initial priming may have also impacted the epitope responses observed.
Serum ELISAs showed negligible differences between experimental and control group antibody titers against the BG505-CH505 + N241 probing antigen throughout the duration of the study (Fig. 2d). However, the experimental group antibody titers had better AMC016 recognition after heterologous boosting at week 24, compared to the control group, even after the control group was heterologously boosted at week 39 (Fig. 2e).
Cross-reactive immunity observed via nsEMPEMCross-linking nsEMPEM of polyclonal sera enables detection of lower abundance antibodies44. Hence, prior to heterologous boosting we attempted this approach to reveal which epitopes might be boosted at weeks 24 and 39. The 2D class averages of samples cross-linked with the heterologous trimer suggests FP-recognizing Abs are present in plasma, which had not been observed in nsEMPEM analysis with BG505-CH505 + N241 trimer (non-crosslinked) (Table S1). This suggestion was corroborated by the FP-ELISA data that showed FP-recognizing pAbs at the week 14 and 28 time points in both groups (Fig. 2c). Thus, FP-directed Abs were elicited at low abundance and/or low affinity levels in the serum by the autologous immunogens. In all the animals, the 2D class averages also show the presence of a base-binding Ab that was able to cross-link with the heterologous immunogen prior to its immune exposure (Table S1). These observations suggest that cross-linked nsEMPEM analysis can reveal cross-linking epitopes between heterologous antigens and can be a valuable tool for vaccine design and boost immunogen selection for real time study evaluation45.
Flow cytometry analysis did not reveal remarkable changes in GC populationsTo gain visibility of the GC dynamics over the course of immunization, longitudinal lymph node fine needle aspirates (FNAs) were performed on the draining inguinal lymph nodes of each animal at several timepoints throughout the study (Fig. S3). Cells collected during the FNAs were stained and analyzed using flow cytometry (Fig. 3). Overall, GC B cell and TFH cell responses were observed in the sampled lymph nodes after initial priming and until after the final boost (Fig. 3a, b). GC response (as measured by GC B cell and TFH cell percentage) was largely unchanged after the boosts at weeks 12 and 24. Beginning at week 24, in addition to surface marker staining, FNA cells were also stained with a fluorescent probe of the heterologous boost immunogen. Interestingly, in the experimental group, the percentage of GC B cells collected from the lymph nodes that are positive for the heterologous AMC016 trimer are greater at week 24 than week 28, four weeks post-heterologous boost (Fig. 3c). Despite no immunization with the AMC016 trimer, the control group exhibited a slight increase in heterologous AMC016 trimer recognition after BG505-CH505 + N241 trimer boosts at week 24 (Fig. 3c).
Fig. 3: Flow cytometry reveals limited changes in GC B and TFH cell populations.
a Quantification of GC B cell kinetics as a percentage of total CD20 + B cells. Experimental group shown in blue circles while control group are shown in black circles. b Quantification of GC Tfh cell kinetics as a percentage of total CD4 + T cells. c % GC B cells that are AMC016+ following the boosts at weeks 24 and 39. Individual animals are shown on the graph to the right with animal RQk18 being the highest binder of the experimental group (depicted as a blue triangle).
Serum neutralization detected after AMC016 heterologous boostingWe assessed heterologous or autologous neutralization of serum samples longitudinally using three pseudovirus (PV) neutralization panels: a PV panel sensitive to FP-targeting bnAbs, a BG505- and a CH505-specific mutant PV panel (Fig. 4a–c, respectively) (Tables S2, S3, and S4, respectively). The former is comprised of ten heterologous PVs selected for their neutralization sensitivity to FP-specific bnAbs10,23. Week 14 serum neutralization analysis revealed sporadic and weak neutralization for the FP sensitive panel, though slightly stronger against 3988.25 Tier 2 PV (Fig. 4a; top panel). There was no detectable neutralization against BG505 and relevant mutant panel, except for two animals, one from each of the groups, that showed weak neutralization against the N611A mutant PV, which does not have a glycan present at position 611 of gp41 (Fig. 4b; top panel). When tested against the CH505 PV panel, there was detectable neutralization against the Tier 2 transmitter/founder (TF) virus and against the Tier 1 A w4.3 virus for the animals in both groups except 1 RM in the control group (Fig. 4c).
Fig. 4: Serum neutralization sporadically observed after heterologous boosting.
a Comparison of FP-Sensitive pseudovirus panel serum neutralization between weeks 14 (top), 28 (middle), and 43 (bottom). (Experimental group is shown in magenta circles and the control group in black circles). Y-axis is in log scale. b Comparison of BG505 pseudovirus panel serum neutralization between weeks 14 (top), 28 (middle), 43 (bottom). Y-axis is in log scale. c Comparison of CH505 pseudovirus panel serum neutralization. Y-axis is in log scale.
Week 28 serum samples were analyzed for neutralization. One animal, RQk18, from the experimental group neutralized six of the nine PVs of the FP sensitive panel, all of which contained FP sequence identical to the FP sequence used in the immunogens of this study (Fig. 4a; middle panel and Table S2). None of the control animals nor the rest of the experimental group developed cross-neutralizing capabilities for the viruses assessed in the FP panel except weak neutralization of Tier 2 3988.25 virus (Fig. 4a; middle panel). Among the control group animals, none of them neutralized the BG505 PVs except weakly against the Tier 2 N611A PV seen in two of the animals (Fig. 4b; middle panel). Within the experimental group, one animal, RQk18, neutralized five of six BG505 PV panel and four out of the six animals neutralized the N611A PV (Fig. 4b; middle panel and Table S3). There was less serum neutralization of CH505 TF PV among the experimental group at this time point compared to the control group, which is expected since the experimental group received AMC016 trimers, and control received the BG505-CH505 + N241 chimeric immunogen at week 24 (Fig. 4c and Table S4).
After the week 39 boost with AMC016 trimer, FP responses were observed via nsEMPEM in both experimental and control groups in five out of six animals (Fig. 2a). At week 43, one animal in the control group weakly neutralized one of nine PVs on the FP sensitive panel (Fig. 4a; bottom panel and Table S2) and five of the six mutant BG505 panel (Fig. 4b; bottom panel and Table S3). After this second AMC016 immunization, two more animals in the experimental group were able to neutralize additional PVs in the BG505 panel (Table S3) but animal RQk18 continued to weakly dominate in the FP sensitive panel neutralization with two other animals showing even weaker neutralization capacity (Table S2).
To test whether the weak serum neutralization profiles observed were mediated through FP recognition, we conducted an FP competition neutralization assay on a subset of PVs, whereby serum samples were titrated with FP-peptide, to capture any FP-recognizing antibodies in the serum (Fig. S4 and Table S5). FP-peptide titration with VRC34.01 positive control showed significant decrease in PV neutralization, however, there was no impact on the serum neutralization profiles from immunized animals. The FP competition neutralization results suggest that the weak serum neutralization observed in a subset of the animals in the larger subset of PVs tested were due to non-FP directed neutralization.
CryoEMPEM analysis reveals high resolution on-target and off-target immune responsesTo better understand the types of FP region responses that were elicited in the animals and overall immunogenicity of our constructs, we conducted cryoEMPEM analysis23,26 on two animals - RQk18 at week 43 and RUu18 at week 14 time points (Fig. S5 and Table S6) - to define the molecular interactions between antigen and pAb. Animal RQk18 developed weak neutralization breadth among the heterologous FP-sensitive PV panel after week 24 boosts, which persisted through the week 39 boosts. CryoEMPEM analysis of RQk18 week 43 pab response complexed with the heterologous AMC016 trimer resulted in three high-resolution maps: 2 define distinct pAb classes targeting the base of the trimer and one shows a pAb bound to the FP (Fig. S5a). Animal RUu18 exhibited what appeared to be an “FP” response via nsEMPEM at week 14 despite FP-ELISA indicating poor FP recognition among the polyclonal IgG responses (Fig. 2c, green triangle and Fig. S2a). Multiple classes of pAbs were classified that targeted off-target BG505-CH505 epitopes: the base, V1V2V3 region and the non-specific “FP” (Fig. S5b).
RQk18 week 43 - FPAnimal RQk18 elicited a heterologous, neutralizing Ab response after week 24 AMC016 trimer boost, as determined by neutralization assays (Fig. 4). To determine whether we could interpret high-resolution structural data and infer sequence information of the bound, polyclonal Abs, we turned to cryoEMPEM, which revealed two epitopes targeted after boosting with the heterologous trimer - the FP and base (Fig. S5a). We resolved a 3.1 Å map with well-resolved density for the FP epitope as well as the variable region of the pAb, RQk-FP-A (Fig. 5a).
Fig. 5: FP-targeting versus FP-proximal immune responses share overlapping epitopes.
a RQk-FP-A pAb1 from week 43 in complex with Heterologous Boost (left panel– side view; right panel – bottom view). The heavy chain (HC) is depicted in orange while the light chain (LC) is shown in yellow. The FP is shown in purple, the N88 glycan in green, gp120 subunit in grey and the gp41 subunit in white. b RQk-FP-A pAb inferred hydrophobic and aromatic residues along its HCDR3, LCDR1 and LCDR3 form a hydrophobic pocket that stabilizes the N-terminus of the FP. c RQk-FP-A pAb interacts with the N88 glycan and C1/C2 beta strands as well as the HR2 domain of gp41. d RUu-FP-1 cryoEM map (Left – bottom view; right – side view). e RUu-FP-1 pAb makes contacts with the C1 beta strand, HR2 and canonical FP glycans N241 and N88.
The structural analysis revealed a fully resolved N-terminus of the FP, stabilized by aromatic side chains on the heavy and light chains of the Ab (Fig. 5b). The HCDR3 of the heavy chain had a high abundance of aromatic residues, which help to stabilize the FP, a highly hydrophobic region (Fig. 5b). This hydrophobic string of aromatic residues in the HCDR3 feature has been observed in two other bnAbs that target the FP - PGT151 and ACS20218,46. Such evidence indicates that NHPs are capable of eliciting FP Abs that have similar features to FP-directed human bnAbs. Apart from the HCDR3, the heavy chain contacts to the trimer include the CDR1 and CDR2 loops and a few residues of the framework region (FWR) 3 region. The light chain helps stabilize the N-terminus of the FP via CDRL3 loop interactions. Contacts with canonical FP Ab as mentioned earlier were also observed for this pAb (Fig. 5c).
RUu18 week 14 - C1/C2 non-specific “FP” adjacent responsesPrior structural analysis of neutralizing, FP-targeting Abs have shown that the N-terminus of the FP is not typically resolved unless stabilized by an Ab10,11,13,23. NsEMPEM suggested animal RUu18 mounted an FP response by week 14 of the study, but an FP-specific ELISA suggested little FP interaction (Fig. S2a and Fig. 2c, green triangle). This indicated that there were immunogenic residues near the FP epitope that could not be resolved from true FP responses at low resolution. At high resolution, the “FP” Ab did not show engagement with the N-terminus of the FP (Fig. 5d). Rather, the first FP residue observed interacting with RUu-FP-1 pAb is L520 (Fig. 5e). The pAb instead predominantly interacted with the following residues: E83 main chain (99.5% global prevalence), I84 (45.21% prevalence), V85 (39.04% global prevalence), E87 (56.06% global prevalence), K231 (81.26% global prevalence), M535 (13.72% global prevalence). Previous epitope analysis of anti-FP bnAbs has shown that these non-FP residues commonly interact with anti-FP Abs23, consistent with the high overlap with the FP epitope despite lack of specific recognition of the FP N-terminus. Although this response was not visualized for the other control group members at week 14, FP-ELISA titers and nsEMPEM of week 43 results for the control group suggests cross-reactivity of C1/C2-partial FP response within all the control groups members and one of the experimental groups (LJ66) (Fig. 2c and Fig. S2).
RUu18 week 14 - V1V2V3Three different V1V2V3-targeting pAb maps were resolved to ~4 Å from cryoEMPEM analysis of RUu18 at week 14 (Fig. S6). Although this Ab response was not observed at the week 4 nsEMPEM analysis in either group, complexing polyclonal fabs isolated from week 14 with the BG505-CH505ΔN241 antigen showed binding of a V1V2V3 Ab to the priming trimer. Glycan analysis of the immunogens revealed an under occupancy of conserved glycans near the variable loops of gp120 of the boosting immunogen used at the week 12 boost for both groups (Fig. 1c). Two of the three maps revealed a hydrophobic pocket that was created by hydrophobic residues in the substituted V1, V2, and V3 loops (Fig.
Comments (0)