
Remember me
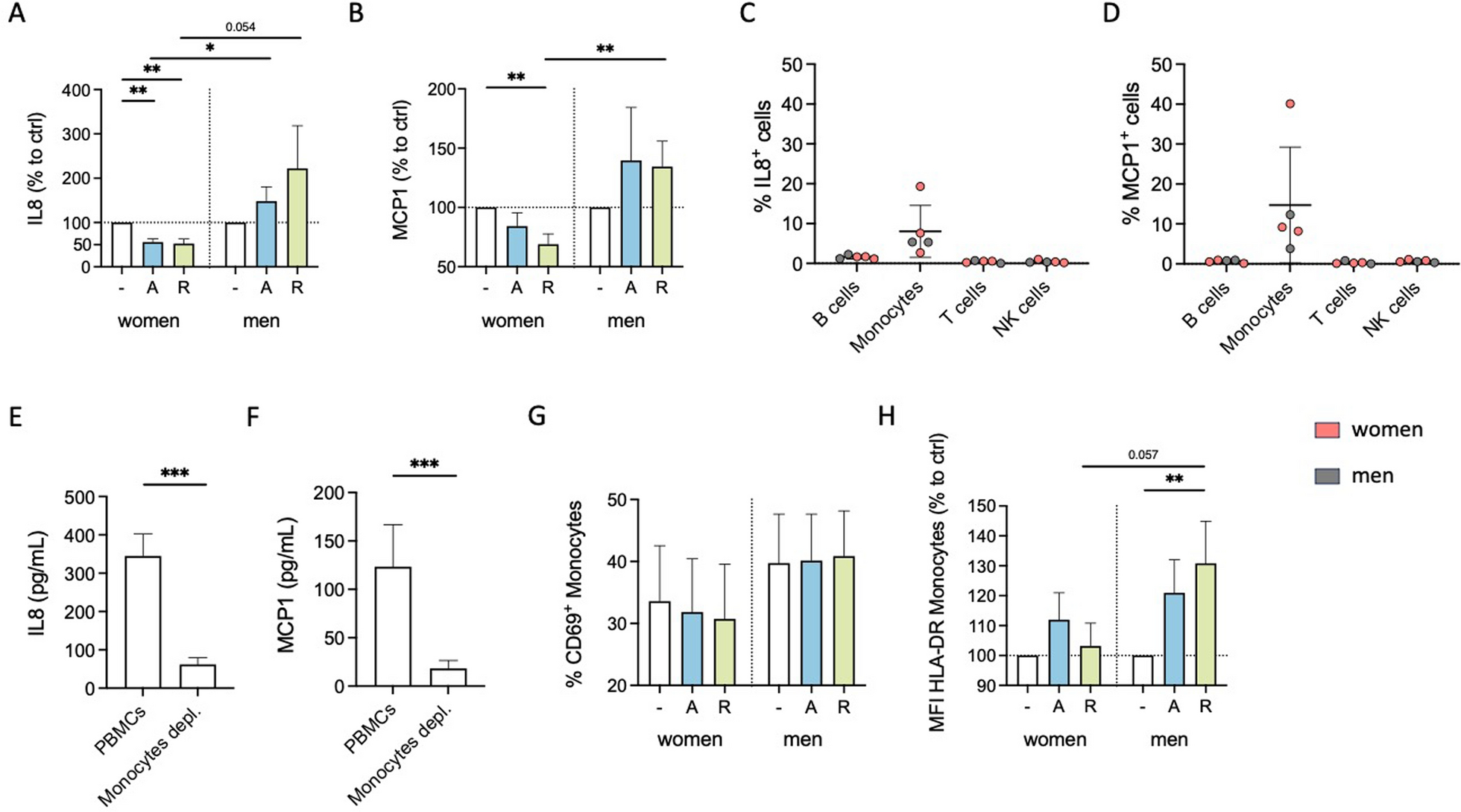


Cytokine secretion is an important immune-modulating property since it leads to the regulation of inflammatory responses and the coordination of cellular communication within the immune system [43, 44]. We investigated whether stimulation with the D1-like agonist A68930 or the D2-like agonist Ropinirole can influence the cytokine secretion of PBMCs from healthy women and men under physiological conditions in vitro. IL8 and MCP1 were detected at measurable levels under physiological conditions (Supplementary Fig. 2A and B), whereas other cytokines were below the detection limit (measured via LEGENDplex™ Human Inflammation Panel 1; data not shown). Upon D1- and D2-like stimulation, there was a significant decrease in IL8 and MCP1 secretion by PBMCs from women. At the same time, this was not observed in men (Fig. 1A and B, Supplementary Fig. 2C and D).
Fig. 1
DR stimulation reduces physiological cytokine secretion and activation marker expression of monocytes from women compared to men. A, B IL8 (A) and MCP1 (B) levels in supernatant from PBMCs of women and men after 24 h in cell culture, with and without in vitro stimulation by A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) measured via ELISA; n = 11–13 per group; normalized to unstimulated control. Basal levels are presented in Supplementary Fig. 2A and B. C, D Percentage of IL8+ (C) and MCP1+ (D) B cells, monocytes, T cells, and NK cells after 24 h in culture without stimulation measured via flow cytometry; n = 5 per subtype. E, F IL8 (E) and MCP1 (F) levels in supernatant from mixed PBMCs and CD14+ monocyte-depleted PBMCs after 24 h in culture without stimulation measured via ELISA; n = 11–13 per condition. G, H Percentage of CD69+ monocytes (G) and expression of HLA-DR on monocytes (H) from women and men after 24 h in culture of mixed PBMCs, with and without stimulation by A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) measured via flow cytometry; n = 13–14 per group; normalized to unstimulated control. Basal levels are presented in Supplementary Fig. 2G and H. Mann–Whitney test was used for comparing unpaired data of women and men, and Wilcoxon test for comparing paired data including unstimulated vs. stimulated samples as well as mixed PBMCs vs. monocytes depleted PBMCs; *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
Previously, we already showed that DRD1, DRD2, DRD3 and DRD4, but not DRD5, are expressed on all PBMC subsets, namely B cells, T cells, NK cells, and monocytes [3], thus, being potentially responsible for the observed changes in cytokine secretion. Intracellular IL8 and MCP1 staining (Fig. 1C and D) and depletion of monocytes from PBMCs showing a strong reduction of these secreted cytokines (Fig. 1E and F) confirmed that their source are the monocytes. Furthermore, DR stimulation itself did not induce IL8 or MCP1 production in B cells, T cells or NK cells (Supplementary Fig. 2E and F).
Additionally, we investigated whether DR stimulation affected the activation status of monocytes, as indicated by the percentage of CD69+ monocytes and the expression of activation markers HLA-DR, CD86, and CD38. While we did not observe a significant impact of dopaminergic stimulation on CD69-expressing monocytes (Fig. 1G) or the expression of CD86 and CD38 (Supplementary Fig. 2I-L) in both sexes, D2-like stimulation resulted in an upregulation of HLA-DR on monocytes from men but not on monocytes from women (Fig. 1H).
B cells from male subjects have a higher expression level of DRD1 and DRD3 than B cells from womenBased on these sex-specific effects of DR agonists on cytokine secretion, we hypothesized that the differences could be attributed to a sex-specific expression of DRs on PBMCs. To test this hypothesis, we analyzed the expression patterns of DRs via flow cytometry. All DRs except DRD5 were found on all PBMC subsets (data not shown). These results are in line with previous findings, except of DRD5, which was described on immune cells in other studies [2, 22]. Consistent with a previous study [3], we did not observe significant differences in the expression of DRs on monocytes between women and men (Fig. 2A). However, higher expression levels of DRD1 and DRD3, but not DRD2 and DRD4, were found in B cells from men compared to women (Fig. 2B).
Fig. 2
Higher DRD1 and DRD3 expression in B cells from men was partly regulated by estradiol and testosterone. A, B Basal expression of DRD1, DRD2, DRD3, and DRD4 on monocytes (A) and B cells (B) from women and men measured via flow cytometry; n = 17–19 per group. C, D DRD1 expression on B cells from women and men after 24 h stimulation with E2 (10–8, 10–9, 10–10 M, C) and DHT (10–7, 10–8, 10–9 M, D) in mixed PBMC culture measured via flow cytometry and normalized to DMSO control; n = 8 per condition. E, F DRD3 expression on B cells from women and men after 24 h stimulation with E2 (10–8, 10–9, 10–10 M, E) and DHT (10–7, 10–8, 10–9 M, F) in mixed PBMC culture measured via flow cytometry and normalized to DMSO control; n = 8 per condition. G, H Correlation of DRD1 expression on B cells from women and men with basal estrogen (G) and testosterone (H) levels in plasma; n = 17–18 per group. I, J Correlation of DRD3 expression on B cells from women and men with basal estrogen (I) and testosterone (J) levels in plasma; n = 17–18 per group. Women: orange; men: grey. Unpaired t-test was used for comparing women and men, while simple linear regression was used to analyze the correlation of DR expression with sex hormone levels. One-way ANOVA with Geisser-Greenhouse correction and Dunnett multiple comparisons test was used for statistical testing of sex hormone receptor (SHR) stimulation with three concentrations of E2 and DHT. Mann–Whitney test compared the effects of SHR stimulation between women and men at the same hormone concentrations; *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
Other studies have already reported that the expression of DRs seems to correlate with the level of sex hormones [31,32,33]. Thus, we stimulated PBMCs with 17β-estradiol (E2) and dihydrotestosterone (DHT) and looked for a change in DRD1 and DRD3 expression on B cells afterwards. We found an upregulation of DRD1 on male compared to female B cells (Fig. 2C and D), however, not for DRD3 (Fig. 2E and F). This upregulation observed only in men is not related to a higher amount of sex hormone receptors on male B cells, since our flow cytometry analysis did not reveal differences in the expression levels of estrogen receptors ERα, ERβ, and GPR30, as well as androgen receptor AR between both sexes (Supplementary Fig. 3A-D).
Next, we measured E2 and testosterone in the plasma to test for a possible correlation with DR expression. Surprisingly, we found comparable levels of E2 in plasma from both women and men in our cohort (Supplementary Fig. 3E). This finding can be partly explained by the lower E2 levels in women taking hormonal contraceptives (26% of the total cohort) compared to those without (Supplementary Fig. 3F). As expected, women in our cohort exhibited significantly lower testosterone levels compared to men (Supplementary Fig. 3G). The direct effect, that was observed after sex hormone stimulation on DRD1 expression, could not be confirmed in correlation analyses between E2 or testosterone levels with DR expression on B cells (Fig. 2G–J), underscoring that the level of sex hormones in plasma cannot be used as a predictor for DR expression on immune cells.
DR stimulation led to a slight but women-specific increase in B cell activationSince we observed differently expressed DRD1 and DRD3 in B cells from women and men, we next investigated whether dopaminergic stimulation influences activation markers on B cells. Interestingly, D2-like stimulation significantly increased CD86 on B cells in women but not men, and the same tendency was observed for D1-like stimulation (p = 0.094; Fig. 3A), suggesting a higher activated phenotype for B cells from women upon dopaminergic stimulation. While CD69 did not show a significant change after DR stimulation (Supplementary Fig. 4D), HLA-DR was slightly upregulated in both sexes, indicating a not-sex-specific effect (Supplementary Fig. 4E).
Fig. 3
DR stimulation increased activation of female B cells. A CD86 expression on B cells from women and men after 24 h in mixed PBMC culture, with and without stimulation by A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) measured via flow cytometry; stimulated samples are normalized to unstimulated control; n = 13–14 per group. Basal levels are presented in Supplementary Fig. 4A. B Representative flow cytometry plots of complete and CD14+ monocyte-depleted PBMCs. C CD86 expression on B cells from women and men after 24 h in monocyte-depleted PBMC culture, with and without stimulation by A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) measured via flow cytometry; stimulated samples are normalized to unstimulated control; n = 7–8 per group. D Gating strategy for B cell subsets (1, naïve B cells: IgD+CD27-; 2, marginal zone-like B cells: IgD+CD27+; switched memory B cells: IgD−CD27+). E CD86 expression on naïve, marginal zone-like, and switched memory B cells from women and men after stimulation of mixed PBMCs with A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) for 24 h normalized to unstimulated control measured via flow cytometry; n = 13–14 per group. Mann–Whitney test was used for testing statistical significance between unpaired data of women and men. Wilcoxon test was used for the comparison of paired data including unstimulated vs. stimulated samples; *p ≤ 0.05, ***p ≤ 0.001
To test whether the increased activation of B cells was due to indirect effects mediated by monocytes, we depleted monocytes from the cell culture (Fig. 3B) and examined the effects of dopaminergic stimulation. D2-like stimulation still caused an upregulation in CD86 expression exclusively in women (Fig. 3C) as observed for B cells in mixed PBMCs, thus excluding a possible secondary effect mediated through monocytes.
Considering that DR expression in B cells has been reported to correlate with different stages of B cell development [3], we analyzed whether the observed differences in CD86 regulation could be attributed to specific B cell subsets. Thus, we stained for IgD+CD27− naïve B cells, IgD+CD27+ marginal zone-like B cells and IgD−CD27+ switched-memory B cells (Fig. 3D). All three B cell subsets showed an increase in CD86 expression exclusively in women following dopaminergic stimulation (Fig. 3E).
These findings provide strong evidence for the sex-specific effects of dopaminergic stimulation on B cells.
B cells contribute to the sex-specific cytokine decrease of female monocytes after DR stimulationNext, we investigated whether the presence of B cells influences the downregulated cytokine secretion by monocytes observed exclusively for women. To explore this, we depleted B cells from the culture of mixed PBMCs (Fig. 4A) and measured the levels of IL8 and MCP1 in supernatant from B cell-depleted fraction from women and men. The downregulation of IL8 and MCP1 secretion from female monocytes observed in mixed PBMC cultures after stimulation with dopaminergic agents (Fig. 1A and B) was not seen after B cell depletion (Fig. 4B and D). A change in IL8 and MCP1 production also by male monocytes was seen in the absence of B cells (Fig. 4C and E), suggesting a possible B cell-dependent mechanism.
Fig. 4
Sex-specific cytokine secretion of monocytes after DR stimulation is dependent of B cells. A Representative flow cytometry plots of mixed PBMCs and CD19+ B cell-depleted PBMCs. B, C IL8 levels in supernatant from PBMCs and B cell-depleted PBMCs from women (B) and men (C) after 24 h in culture, with and without stimulation by A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) measured via ELISA; normalized to unstimulated control; n = 8–9 per group. D, E MCP1 levels in supernatant from PBMCs and B cell-depleted PBMCs from women (D) and men (E) after 24 h in culture, with and without stimulation by A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) measured via ELISA; normalized to unstimulated control; n = 8 per group. F, G Diagrams illustrating the interplay between B cells and monocytes after DR stimulation, showing effects on activation markers and cytokine secretion for cells from women (F) and men (G); created in BioRender.com. Wilcoxon test was used for paired data comparisons including PBMCs vs. B cells depleted as well as unstimulated vs. stimulated samples. Mann–Whitney test was used for testing statistical significance between unpaired data of women and men; *p ≤ 0.05
In summary, DR stimulation leads to increased activation of female B cells, which can subsequently induce a reduction in the activation and cytokine secretion of monocytes. Conversely, this activation in B cells in response to DR stimulation is not observed for men, and consequently, there is no downregulation in activation and cytokine secretion of monocytes (Fig. 4F and G).
Acute stimulation induces switch to proinflammatory effect of DR agonists on cytokine secretion in womenIn a previous study, we demonstrated sex-specific responses to DR stimulation in patients with rheumatoid arthritis (RA), a chronic inflammatory disease [3]. Additionally, changes in dopaminergic effects due to the coactivation of the MAPK signaling pathway are reported [45]. To elucidate whether sex-specific differences can also be observed under acute inflammatory conditions, we introduced CpG, which also leads to activated MAPK signaling [46]. We demonstrated that the stimulation with CpG activated both B cells and monocytes without indirect B cell influence (Supplementary Fig. 5A and B).
Our findings showed that, in the presence of CpG, IL8 was still mainly produced by monocytes as shown by the intracellular staining of IL8 (Supplementary Fig. 6A) and the reduction in IL8 level after depletion of monocytes from PBMCs (Supplementary Fig. 6B). However, a CpG-induced increase in IL8 expression has also been found for B cells (Supplementary Fig. 6A). With B cells and monocytes as mixed sources for IL8, we observed a comparable increase in IL8 by CpG stimulation in both sexes, with no effect by dopaminergic stimulation, unlike under physiological conditions, which presented monocytes as the exclusive IL8 source (Supplementary Fig. 6C and D).
We focused on the secretion of MCP1 since only monocytes and no other PBMC subset showed a detectable level of MCP1-expressing cells, which were upregulated by CpG stimulation after 24 h (Fig. 5A, Supplementary Fig. 6E). Furthermore, the depletion of monocytes resulted in nearly undetectable levels of MCP1 (Fig. 5B), underscoring that monocytes are the sole source of MCP1. Interestingly, CpG stimulation increased MCP1 expression in both sexes; however, combined with dopaminergic agents, an even more pronounced increase was observed only for women but not men (Fig. 5C). Thus, dopaminergic stimulation under inflammatory conditions switched the anti-inflammatory phenotype to a proinflammatory effect regarding MCP1 secretion only in women (Supplementary Fig. 6F and G).
Fig. 5
Inflammatory condition turned DR stimulation-induced characteristics of female monocytes into proinflammatory phenotype. A Percentage of MCP1+ B cells, monocytes, T cells, and NK cells after 24 h in mixed PBMC culture, with or without CpG (0.195 μM) stimulation, measured via flow cytometry; n = 12 per condition. B MCP1 levels in supernatant from mixed and monocyte-depleted PBMCs after 24 h of CpG stimulation measured via ELISA; n = 14 per condition. C MCP1 levels in supernatant from PBMCs of women and men after 24 h in culture with CpG (0.195 μM) with or without A68930 (A, 10–7 M) or Ropinirole (R, 10–6 M) measured via ELISA; normalized to CpG control; n = 11–13 per group. Basal levels of unstimulated and CpG stimulated samples are presented in Supplementary Fig. 5C. D-G Percentage of CD69+ monocytes (D) and expression of HLA-DR (E), CD86 (F) and CD38 (G) on monocytes from women and men after 24 h in culture of PBMCs with or without CpG (0.195 μM) stimulation and after stimulation with CpG + A68930 (A, 10–7 M) and CpG + Ropinirole (R, 10–6 M) measured via flow cytometry; normalized to CpG control; n = 14 per group. Basal levels of unstimulated and CpG stimulated samples are presented in Supplementary Fig. 5D-G. Wilcoxon test was used for paired data comparisons including samples of two different stimulations as well as PBMCs vs. monocytes depleted. Mann–Whitney test was used for testing statistical significance between unpaired data of women and men; *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
In addition to MCP1, IL1β and IL18 also appeared to be monocyte-specific since the depletion of monocytes led to an almost complete loss of these cytokines in the supernatant (Supplementary Fig. 6H and I). The release of both cytokines was induced by CpG (Supplementary Fig. 6J and K), confirming its activating effects also on monocytes, and was further increased by DR stimulation in women (Supplementary Fig. 6L and M). This supportsthe proinflammatory phenotype of female monocytes due to DR stimulants under these inflammatory conditions. This finding was further strengthened by an increase in CD69+ monocytes and the increased expression of activation markers HLA-DR and CD86 on monocytes induced by DR stimulation, which was not statistically significant in men (Fig. 5D–G). Notably, the switch in the effects from physiological to inflammatory conditions was pronounced for the number of CD69-expressing monocytes, but similar for both women and men (Supplementary Fig. 7A and B). This was also evident for CD86 and CD38, resulting in a stronger upregulation after DR stimulation in the presence of CpG than without additional stimulus (Supplementary Fig. 7E–H), but not for HLA-DR (Supplementary Fig. 7C and D).
In summary, our results indicate that the effects of dopaminergic stimulation are shifted to a proinflammatory response regarding cytokine secretion and monocyte activation by an acute inflammatory stimulus specifically in women.
Interestingly, DR stimulation also led to an increase in HLA-DR and CD86 on T cells under physiological as well as inflammatory conditions (Supplementary Fig. 8) suggesting an increased activation for this PBMC subtype. However, these effects on T cells were independent of sex and the presence of an acute inflammatory stimulus, presenting the observed responses of B cells and monocytes to be cell specific.
Monocytes exhibit a B cell-independent proinflammatory phenotype after DR stimulation under inflammatory conditionsTo investigate whether the influences of dopaminergic agonists on monocyte activation and cytokine production are still driven by B cells also under inflammatory conditions, we depleted the B cells from the mixed PBMCs and analyzed MCP1 and activation marker expression after dopaminergic stimulation. Interestingly, the upregulation of MCP1 for women following DR stimulation was not observed after B cells depletion (Fig. 6A). However, the upregulation of activation markers in women and men seemed to be B cell-independent (Fig. 6C–J). Thus, we hypothesized that the B cells do slightly support the upregulation in MCP1 by monocytes under inflammatory conditions, but that DR stimulation also has a direct effect in CpG-activated monocytes leading to an even more pronounced proinflammatory phenotype.
Fig. 6
Proinflammatory phenotype of monocytes from women after DR stimulation is independent of B cells under inflammatory condition. A, B MCP1 levels in supernatant from mixed and B cell-depleted PBMCs from women (A) and men (B) after stimulation with CpG (0.195 μM), CpG + A68930 (A, 10–7 M) or CpG + Ropinirole (R, 10–6 M) measured via ELISA; normalized to CpG control; n = 7–8 per group. C, D Percentage of CD69+ monocytes from women (C) and men (D) after stimulation with CpG (0.195 μM), CpG + A68930 (A, 10–7 M) or CpG + Ropinirole (R, 10–6 M) measured via flow cytometry; normalized to CpG control; n = 7 per group. E–J HLA-DR (E, F), CD86 (G, H), and CD38 (I, J) expression on monocytes from women (E, G, I) and men (F, H, J) after stimulation with CpG (0.195 μM), CpG + A68930 (A, 10–7 M) or CpG + Ropinirole (R, 10–6 M) measured via flow cytometry; normalized to CpG control; n = 7 per group. Wilcoxon test was used for paired data comparisons including unstimulated vs. stimulated samples as well as PBMCs vs. B cells depleted; *p ≤ 0.05
Our findings about the effects of DR stimulation on PBMCs from women and men under physiological and acute inflammatory conditions are summarized in Fig. 7. For women, DR stimulation of B cells leads to an upregulation of the surface marker CD86 under healthy conditions (Step 1). Following this activation, B cells affect monocytes (Step 2), which results in the downregulation of the proinflammatory cytokines MCP1 and IL8, thus leading to an anti-inflammatory response (Step 3). Contrary for men, CD86 on B cells is not affected by DR stimulation, correlating with no downregulation of MCP1 and IL8. These effects highlight sex-specific differences in B cell-mediated immune modulation after DR stimulation. Under acute inflammatory conditions induced by CpG, which was shown to activate the monocytes directly, the effects of DR stimulation shift to a more proinflammatory phenotype only in women shown by an increased expression of activation markers on monocytes and a higher release of monocyte-derived MCP1. This switch from anti- to proinflammatory effects induced by DR stimulation is less pronounced in men. Concluding, this illustration shows the complexity of immune responses induced by DR stimulation, revealing a dependency of cell-type, activation status and sex.
Fig. 7
Effects of DR stimulation on female and male monocytes under physiological and acute inflammatory condition. DR stimulation dampens cytokine secretion of monocytes under physiological conditions (top), dependent on activated B cells. In men, this effect could not be observed. In acute inflammatory conditions induced by CpG (bottom), DR stimulation induces a switch into a proinflammatory phenotype of monocytes from women but not from men, independent of B cells; created in BioRender.com
Comments (0)