
Remember me



In an effort to further improve purity and efficacy of sEVSKM isolation and characterization, we first sought to identify another potential muscle-specific surface marker besides SGCA. We selected muscle-specific kinase (MuSK), a neuromuscular junction protein, based upon its known high expression in muscle as reported by human protein atlas [40] (https://www.proteinatlas.org/w.proteinatlas.org/) and a prior report suggesting MuSK expression in sEV [41]. We utilized the characterization group (age 16 to < 25 years old) for the isolation of total sEV (n = 6). From total sEV, sEVSKM were isolated using either SGCA (sEVSGCA) or MuSK (sEVMuSK), or both antibodies in combination (sEVSG/Mu) by the immune-isolation method (briefly outlined in Fig. 1A).
Fig. 1
Characterization of sEVSKM isolated from the serum of vervets. A Diagrammatic representation of sEVSKM isolation methodology. Total sEV and sEVSKM were isolated from the serum of monkeys (16 to > 25 years old) and characterized by multiple techniques for their purity. B Line graph representing size and concentration of sEVSKM (n = 6). C–E Representative images showing the structure of sEVSKM (C), surface expression of SGCA (smaller gold particles—marked by red arrows) and CD63 (larger gold particles—marked by black arrow) (D), surface expression of MuSK (smaller gold particles—marked by red arrows) and CD63 (larger gold particles—marked by black arrow) (E) by immunogold labeling coupled with transmission electron microscopy. Images were captured at 49,000 × magnification (n = 3 each), and scale bar is presented at the bottom of each image. F Immunoblot array showing the expression of SGCA and CD63 loaded in sEVSKM (n = 1). G, H. Representative images and bar graphs showing expression of SGCA (G) and MuSK (H) along with CD63 and CD9 by super-resolution microscopy (n = 3). The error bars in the graphical data represent the mean ± SD. I Confocal images captured at 20 × , showing expression of SGCA along with CD63 and MuSK on the sEV immobilized on the agarose beads (n = 3). J Scatter plot and bar graph representing the % abundance of CD63, CD9, CD81, SCGA, MuSK, and GFAP on sEV.SKM surface (n = 5–6)
We first analyzed the total concentration of these sEV by NTA. Interestingly, we observed that SGCA tended to provide a better yield of sEVSKM compared to either MuSK or a combination of both SGCA and MuSK (Supplementary Fig. 1A), though this difference was not statistically significant. Further, we used flow cytometry to assess the surface expression of CD63, a known exosome-related tetraspanin biomarker, on these sEV. Notably, we did not observe any statistically significant difference in CD63 expression between all the three approaches (Supplementary Fig. 1B). Taken together, our data suggested that SGCA alone allows a comparable yield of SEVSKM from biofluid without compromising the purity. Furthermore, sEVSGCA also showed expression of MuSK on their surface, as confirmed using the immunogold method (Supplementary Fig. 1C), which suggested that, although MuSK inclusion did not further improve the yield or purity of sEVSKM, it could serve as a secondary validation marker to assess the purity of isolated sEVSKM. Since we did not observe any advantage of using multiple markers to isolate sEVSKM in terms of yield and purity, a single marker SGCA was used for the isolation of sEVSKM.
While SGCA-positive sEV in circulation have been previously reported [23,24,25,26], a few studies using less sensitive western blot-based methods failed to detect SGCA-positive sEV in the plasma/serum total sEV pool [42, 43]. In this study, we performed extensive characterization of sEVSKM using multiple high-sensitivity methods (Fig. 1). After enrichment from total sEV, utilizing an immune-isolation method using SGCA antibody, sEVSKM were first characterized for their size and concentration using NTA which confirmed that their average size (133.81 ± 2.48 nm) is in the acceptable size limit for sEV (≤ 200 nm) (Fig. 1B). Next, transmission electron microscopy (TEM) confirmed the double membrane-bound structure with a typical size of sEV (Fig. 1C). We then utilized a novel approach of double labeling of sEV with two different-sized gold particle-tagged antibodies (10 nm or 20 nm) and confirmed the surface co-expression of SGCA (Fig. 1D) and MuSK (Fig. 1E) with CD63. Next, we used antibody array to validate the presence of SGCA along with CD63 loaded in sEVSKM (Fig. 1F).
Super-resolution microscopy is a powerful microscopy tool which allows imaging sEV at a single molecular level. Super-resolution microscopy also confirmed the presence of the SGCA and MuSK on the sEVSKM along with exosomal markers CD63 and CD9 (Fig. 1G and H). Interestingly, we observed most of the sEVSKM were positive for SGCA, CD63, and CD9 (Fig. 1G, bar graph) and MuSK, CD63, and CD9 (Fig. 1H, bar graph), confirming the purity and muscle specificity of these vesicles.
Immunofluorescence staining of sEV immobilized on agarose beads, using fluorescent antibodies against CD63 and SGCA, simultaneously confirmed co-localization of CD63 and SGCA on sEVSKM (Fig. 1I; top panel). Similarly, surface co-expression of SGCA with a second muscle-specific marker MuSK on the beads was also confirmed (Fig. 1I; bottom panel). Next, nano-flow cytometry also showed the abundance of skeletal muscle marker SGCA and MuSK along with exosomal surface markers (CD63, CD9, and CD81) on sEVSKM (Fig. 1J). Since total sEV is a heterogenous pool of various cell/tissue-derived sEV, thus the possibility of contamination of other cell/tissue-specific sEV while pulling down a specific population is likely. Thus, we also checked the presence of GFAP-positive sEV (an astrocyte-derived sEV marker) in sEVSKM as a negative marker. Importantly, the negligible presence of GFAP on sEVSKM further confirmed the purity of isolated sEVSKM (Fig. 1J). Altogether, our data suggested that skeletal muscle-derived sEV are present in serum and can be isolated from the blood using SGCA with high purity.
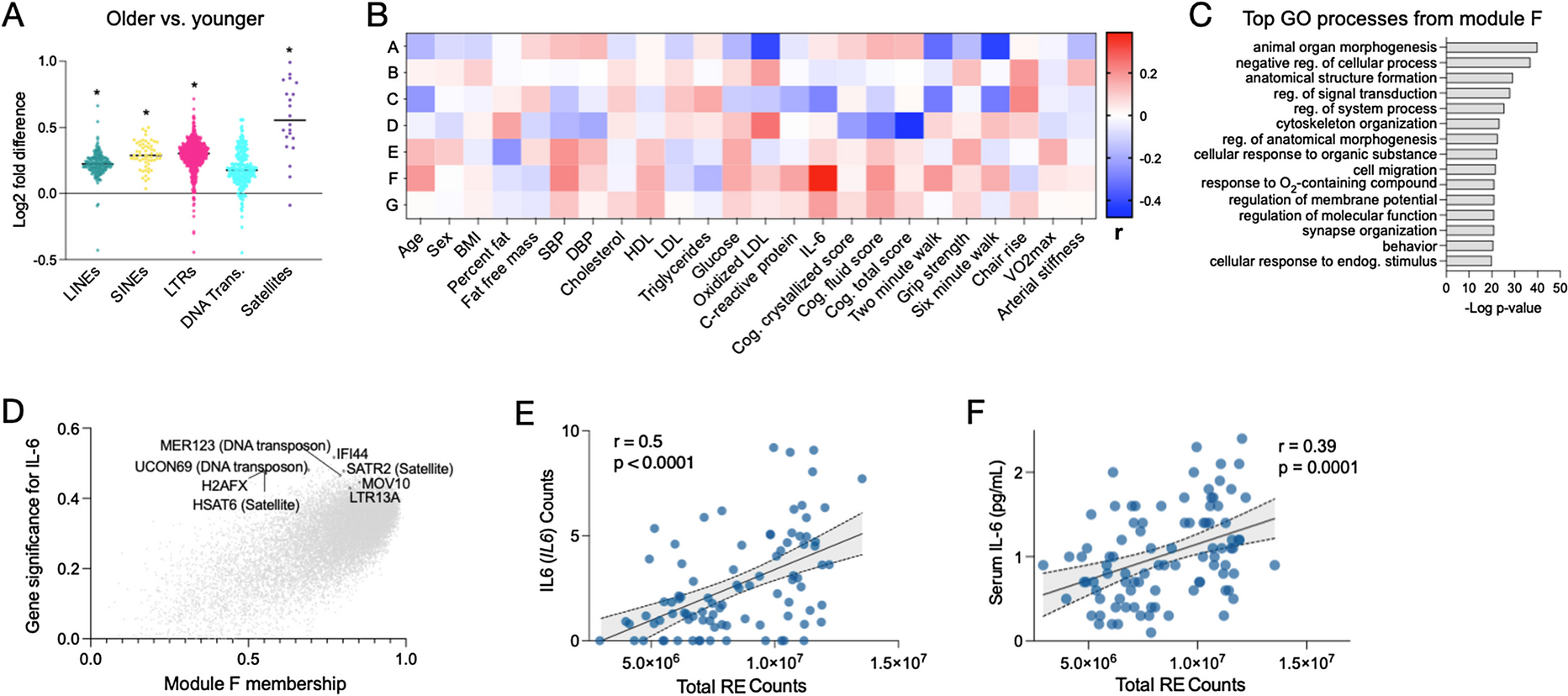
Characterization of sEVSKM for age-associated pathology in young and old vervet monkeysTo establish the translation significance of serum-derived sEVSKM, we isolated total sEV followed by sEVSKM separation from the serum of young (11–15 years; n = 7) and old (25–29 years; n = 11) vervet monkeys following the method described above (Fig. 1A). We first characterized the total sEV by analyzing changes in their secretion profile employing NTA and did not find any significant age-related differences in the total concentration or size of total sEV (Fig. 2A). We next characterized total sEV for percent abundance of SGCA positive sEV (sEVSKM) between young and old vervets using flow cytometry. We observed significant decrease in sEVSKM in the old vervets (Fig. 2B). Since it is well established that muscle mass decreases with age, we speculate that this reduction in the sEVSKM is likely attributable to the overall loss of muscle mass. However, since sEV from muscle cells are also known to promote the differentiation and proliferation of myoblasts [44, 45], it is also plausible that a decrease in sEVSKM secretion may play an active role in exacerbating muscle loss, highlighting the potential of sEVSKM as both, a marker and modulator of muscle health.
Fig. 2
Characterization of serum sEVSKM in young and old vervets. Total sEV and sEVSKM were isolated from the serum of young (≤ 15 years) and old (≥ 25 years) vervets and analyzed. A Bar graphs represent the average concentration and size of total sEV. B Representative scatter plots and bar graphs showing the percentage abundance (mean ± SEM) of SGCA positive sEV in total sEV of young and old vervet. C Bar graphs representing the average sEVSKM concentration (left panel) and size (right panel) for young and old vervets. D, E Bar graphs representing the total protein (D) and RNA (E) content in sEVSKM of young and old vervets (n = 7–11). *p ≤ 0.05
Next, we isolated sEVSKM from the total sEV and characterized them for the concentration and size using NTA. Even though we observed a significantly lower SGCA-positive population in total sEV in old vervets by flow cytometry (Fig. 2B), we did not see a significant difference in size or concentration of sEVSKM after immune-isolation (Fig. 2C), possibly due to the known limited efficiency of the immune-isolation method.
We further analyzed the protein and total RNA cargo of sEVSKM between young and old vervets (Fig. 2D). We observed a significant decrease in protein cargo in old vervets compared to young, while total RNA content remains unchanged (Fig. 2D). Taken together, these findings suggest that aging disrupts not only the secretion of sEVSKM secretion but could also impact their cargo composition.
Characterization of sEVSKM protein cargo for age-associated pathology in young and old vervetsOxidative stress is one of the key underlying factors in age-associated muscle mass/function decline [46, 47]. Due to increased oxidative stress, reactive oxygen species (ROS) accumulation causes protein oxidation leading to subsequent muscle loss. Thus, we next measure the oxidized protein content in sEVSKM. To measure total reversible oxidation (TRO), we employed cysteine-reactive phosphate tag (CPT) to enrich peptides containing cysteine with a wide range of oxidation states including sulfenylation, intra-, and inter-molecular disulfide, glutathionylation, hydropersulfidation, and S-nitrosation, and peptides were identified by LC–MS/MS analysis [48]. We identified 351 and 261 oxidized proteins in sEVSKM of young and old monkeys, respectively (Fig. 3A) representing a decrease in the number of oxidized proteins in the sEVSKM from old vervets. With aging, protein diversity could change due to altered rate of protein translation or disrupted protein homeostasis processes such as protein folding and/or protein complexes formation [49]. In concord with these age-related changes, we detected a similar trend in sEVSKM, both at total protein concentration (Fig. 2D) and oxidized proteins number (Fig. 3A). These findings suggest that sEVSKM cargo could reflect the broader age-related changes in muscle cell.
Fig. 3
Characterization of oxidized proteins in sEVSKM from young and old vervets. A Venn diagram representing the number of oxidized proteins identified in sEVSKM of young and old vervets (n = 3/group, 6 samples were randomly pooled into 3 separate samples). B Volcano plot representing the significantly perturbed oxidized proteins in sEVSKM of young and old vervets. C Bubble plot representing KEGG pathways associated with the significantly perturbed (STN ≤ − 0.3 and ≥ 0.3) oxidized proteins in sEVSKM of young and old vervets. D Bubble plot representing KEGG pathways associated with the oxidized proteins uniquely present in sEVSKM of young vervets
Next, we identified significantly perturbed proteins in sEVSKM from young and old vervets (Fig. 3B). We observed that cofilin 1 (CFL1) and actin-related protein 3 (ACTR3) were the most upregulated oxidized proteins in sEVSKM of old-aged monkeys compared to young-aged monkeys. Both proteins are part of the actin polymerization/depolymerization complex which regulates muscle cell contraction and force generation [50, 51] suggesting a shift in cytoskeletal dynamics with aging. Furthermore, we observed catenin beta 1 (CTNNB1), NOTCH3, heat shock protein 90 alpha class B (HSP90AB1), alpha-2-HS-glycoprotein (AHSG), and eukaryotic elongation factor 2 (EEF2) were significantly downregulated oxidized proteins in sEVSKM of old-aged monkeys compared to their young counterpart. CTNNB1 and AHSG regulate the glucose uptake in muscle cells [52, 53], thus facilitating the muscle energy metabolism. EEF2 and HSP90AB1 play important roles in protein translation and folding processes [54], and NOTCH3 is reported to regulate muscle proliferation [55] contributing to muscle growth and regeneration, all essential for preserving muscle mass. These findings indicate that aging is associated with disruptions in pathways related to muscle metabolism, actin cytoskeleton dynamics, and protein homeostasis, all of which likely contribute to age-related muscle loss. In this regard, Day et. al. recently evaluated biopsies of vastus lateralis muscle in the participants of Study of Muscle, Mobility and Aging (SOMMA) and reported that increased oxidation of key proteins involved in muscle structure and contraction was negatively correlated with the muscle function [56]. Specifically, increased oxidized levels of sarcomeric proteins myomesin-1, myomesin-2, and nebulin were associated with slower walking speed, and increased oxidation of myomesin-2, alpha-actinin-2, and skeletal muscle alpha-actin were associated with lower leg power and strength [56]. This study confirmed that levels of these oxidized proteins in muscle were directly associated with compromised muscle health [56]. In the present study, there was a relatively lower prevalence of oxidized proteins in sEVSKM in old monkeys, perhaps due to the retention of oxidized proteins within the muscle of older monkeys. Taken together, these findings suggest that sEVSKM could play a critical role in the clearance of oxidized proteins within the muscle, thereby protecting the cells from oxidative damage. However, aging may impair this clearance mechanism, leading to the accumulation of oxidized proteins, causing muscle damage. It is important to determine if there is an inverse relationship between oxidized protein content within the muscle vs. exported in the sEVSKM, and ongoing and future studies will provide answers to that and other critical questions.
Next, we analyzed 247 shared proteins in sEVSKM from young and old vervets and identified the significantly altered proteins (STN ≤ − 0.3 and ≥ 0.3). KEGG pathways associated with these perturbed proteins were enriched by the DAVID online tool. We observed several pathways including metabolic pathway, regulation of actin cytoskeleton, cytoskeleton in muscle cells, Rap 1 signaling pathway, biosynthesis of amino acids, HIF-1 signaling, and pyruvate metabolism as the top-hit pathways. In our analysis for oxidized proteins, we found 104 proteins which were detected only in sEVSKM from young vervets. KEGG pathway enrichment of these proteins showed several pathways including metabolic pathway, thyroid hormone signaling, cGMP-PKG signaling, nucleotide metabolism, glutathione metabolism, fatty acid degradation, and one carbon pool by folate (Fig. 3D).
These findings indicate that aging could perturb core metabolic and cytoskeletal regulatory pathways as well as critical signaling and metabolic functions. These age-related alterations in sEVSKM proteins present as potential novel targets for diagnostic as well as therapeutic intervention for mitigating muscle mass/function loss in the elderly. However, further studies are needed to validate these potential targets in corresponding muscle tissues.
Characterization of sEVSKM miRNA cargo for age-associated pathology in young and old vervetsWe next analyzed the miRNA cargo loaded in the sEVSKM of young and old vervets and identified 112 and 88 miRNAs in sEVSKM of young and old vervets, respectively (Fig. 4A). Similar to protein cargo, here also we observed low diversity of miRNA in sEVSKM from old vervets. The biological pathways associated with the most differentially expressed miRNAs were identified using miEAA and ShinyGO tools. Analysis of KEGG pathways associated with the miRNA targets showed that top enriched pathways included focal adhesion, cellular senescence, apoptosis, neurotrophin signaling, cell cycle, relaxin signaling, endocrine resistance, HIF-1 signaling, TNF signaling pathway, T-cell receptor pathways, and adherence junctions (Fig. 4B). Further enrichment for biological processes showed that the most significantly associated biological events were related to metabolic regulation, stress response, and cell death (Fig. 4C).
Fig. 4
Characterization of miRNAs in sEVSKM from young and old vervets. A Venn diagram representing the number of miRNAs identified in sEVSKM from young and old vervets (n = 3/group, 6 samples were randomly pooled into 3 separate samples). B, C. Bubble plot representing the top significantly enriched KEGG pathways (B) and Gene ontology-Biological process (C), associated with perturbed miRNAs
Both oxidized protein and miRNA analyses of sEVSKM consistently identified the involvement of three primary biological processes: metabolic pathways, cell–matrix interactions (actin cytoskeleton, muscle cytoskeleton, adherence junction, focal adhesion), and inflammatory responses. These pathways are known to be involved in age-related muscle mass and function loss. In aging, metabolic changes are considered to be a significant contributor to muscle mass loss [57, 58]. These changes include imbalances in protein synthesis/degradation and impaired glucose metabolism. Skeletal muscle is a key site for glucose uptake where it is actively converted into glycogen [59] serving as a local energy substrate during exercise. Metabolically, muscle is a very flexible organ which rapidly adapts to use glucose, fatty acid, or amino acids as their substrate. Flexibility of skeletal muscle to use lipid or carbohydrate as substrate is associated with different panels of structural proteins [60]. Mitochondrial dynamics also play a critical role in metabolic adaptation [61, 62]. In this study, we found significant enrichment of HIF-1 signaling at both the protein and miRNA levels. HIF 1, a master intracellular oxygen sensor, regulates mitochondrial biogenesis, respiratory chain, TCA cycle, and membrane potential [63]. Further, low-grade persistent chronic inflammation is a commonly observed aging phenotype. The presence of damaged macromolecules, cellular and immune senescence, and increased activation of the coagulation system are some of the possible sources/mechanisms of chronic inflammation in the aging population [64, 65]. Inflammation induces muscle function/mass loss by promoting protein degradation, inhibiting protein synthesis, and activation of satellite cells [66, 67]. Importantly, in peripheral circulation, sEVSKM cargo provided a window into these major metabolic, cytoskeleton, and inflammation-related molecular changes in the skeletal muscle of old vervets.
This study delineates significant advancement in skeletal muscle-derived sEV characterization, including the isolation and purity validation of sEVSKM, utilization of an excellent non-human primate model for aging (a close phylogenic neighbor of humans), and identification of molecular changes with aging. Limitations of this study include the need to validate identified molecular changes in corresponding muscle tissues and the observed molecular changes in sEVSKM by other methods and in multiple possible models. Despite these weaknesses, the present study is a major advancement in the isolation of sEVSKM from serum, validating their purity by multiple methods and identifying potential molecular changes in muscle with aging. Ongoing studies are evaluating these alterations in blood sEVSKM in relation to skeletal muscle biopsy and physical and functional phenotypes in these same subjects.
Comments (0)