
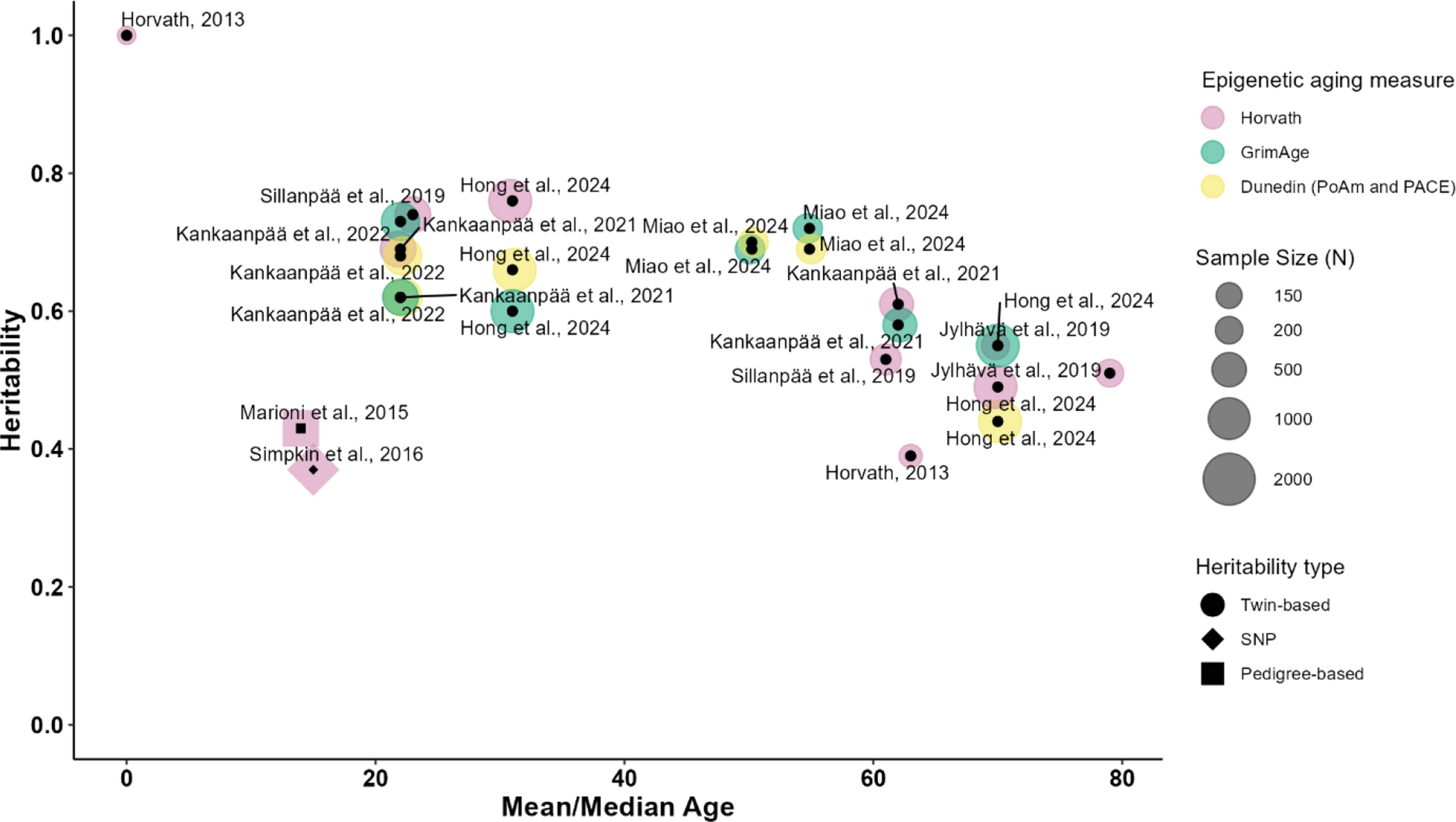
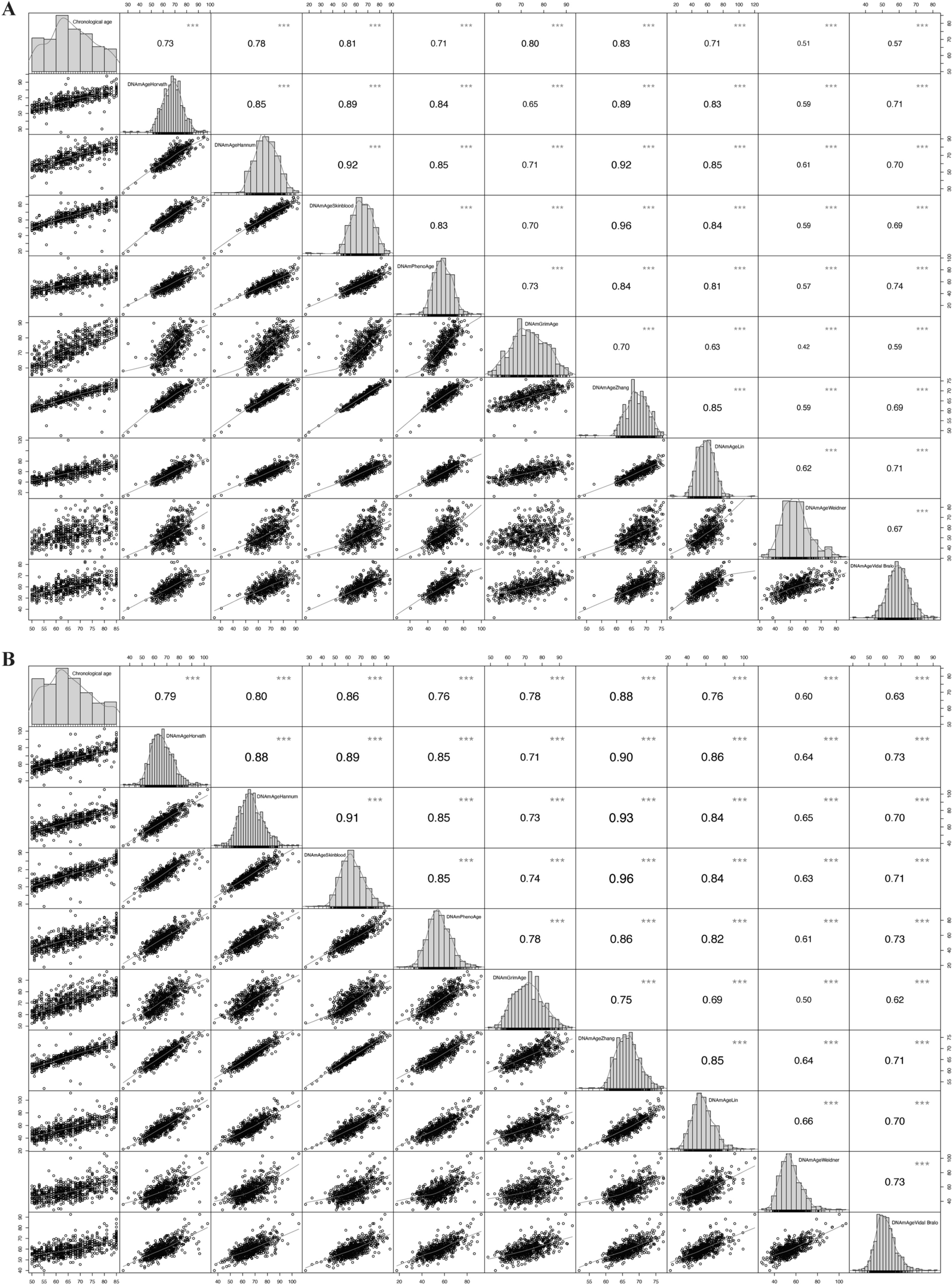
Remember me






We first collected liver cancer and paired normal tissue samples from four patients and performed ATAC-seq sequencing (Fig. S1A). The fragment size distribution of the ATAC-seq samples showed that most insert fragments were relatively small, representing the open chromatin regions between nucleosomes (Fig. S1B). Figure S1C, D demonstrated the enrichment of ATAC-seq signals mainly around transcription start sites (TSSs), which is consistent with previous research, indicating the reliability of sequencing quality. We then annotated these open chromatin regions, revealing that most of the peaks were in promoter and intergenic regions, with a smaller proportion in intronic regions (Fig. S1E), which suggest that transcription factors tend to bind to the promoter regions around TSSs. Subsequently, we used the MAnorm2 algorithm to identify the DARs between liver cancer and normal tissues. We categorized the peaks into those enriched in cancer, enriched in normal, and with no significant difference. (The significance of the differences was calculated via M-values) (Fig. S2A–D).
Genomic characterization and enrichment analysis of differential accessible chromatin regionsTo study the gene regulatory differences between tumor and normal tissues, we performed genomic characterization and enrichment analysis of the DARs. We defined peaks with M-value > 0.5 or < − 0.5 as significantly differential, and obtained 605 peaks significantly enriched in the cancer samples (termed "up") and 400 peaks significantly enriched in the normal samples (termed "down"). Genomic annotation showed that regardless of whether the peaks were enriched in cancer or normal tissues, they were mainly distributed in promoter and distal intergenic regions, with fewer in intronic regions (Fig. 1A), which suggests that the proximal regulatory elements, such as promoters, have higher transcriptional activity in both liver cancer and normal tissues. To further analyze the functions of these DARs, we annotated them to their regional genes and performed enrichment analysis. KEGG pathway enrichment analysis showed that the peaks enriched in cancer samples were mainly associated with genes involved in cell–cell adhesion, HCC, gastric cancer, colorectal cancer, MAPK signaling, and FoxO signaling pathways, while the peaks enriched in normal samples were mainly associated with genes involved in TGF-β, cAMP, and p53 signaling pathways (Fig. 1B). To identify the key transcription factors (TFs) driving the expression differences between cancer and normal tissues, we used HOMER to scan the ± 200 bp flanking regions of the ATAC-seq peaks in the DARs for TF motifs and TF binding sites (TFBS). We found that the DARs enriched in tumor tissues were primarily associated with transcription factor families such as the ETS family, including ETS1, PU.1, Etv2, and Elf4. Additionally, we found that members of the activator protein-1 (AP-1) family, including Jun (JUN-AP1) and Fos (Fosl2), were among the top predicted motifs in the cancer-specific hyper-accessible regions. In contrast, the DARs enriched in normal tissues were mainly associated with CTCF and its paralog BORIS (Fig. 1C). This indicated that changes in chromatin accessibility affected the gene expression of many components of signaling cascades and regulators, and played a critical role in HCC progression.
Fig. 1
Identification and analysis of differential chromatin accessible regions. A Pie charts showing the proportion of up-regulated (left) and down-regulated (right) chromatin accessible regions within the indicated genomic regions in hepatocellular carcinoma(HCC), compared with adjacent normal tissue. B The KEGG pathways significantly enriched by genes in or nearby the up-regulated (left) and down-regulated (right) chromatin accessible regions in HCC. C The top 15 known TF motifs enriched in the up-regulated (left) and down-regulated (right) chromatin accessible regions in HCC
The DARs were positively correlated with the nearest DEGsTo identify DARs potentially driving DEGs, we retrieved matched ATAC-seq and transcriptomic data for liver cancer from TCGA. Leveraging genomic annotation files, we observed that the open chromatin regions within the TCGA-LIHC-ATAC-seq data primarily resided in promoter regions (Fig. S3A). This finding indicated high consistency between the database's genomic distribution characteristics and our sequencing data. Differential analysis of the TCGA LIHC transcriptomic data revealed a total of 1853 DEGs, with 1219 upregulated and 634 downregulated genes (Fig. S3B). By comparing our previously generated DARs with the TCGA-LIHC-ATAC-seq peaks, we identified four sets of overlapping regions: O1 (731 peaks), O2 (559 peaks), O3 (691 peaks), and O4 (1138 peaks) (Fig. S3C). To further investigate potential correlations between DARs and DEGs, we assigned DARs in O1–O4 to their nearest genes. Integration with RNA-seq data revealed 432 and 296 DEGs associated with hyper-accessible regions in tumor and normal tissues, respectively. Notably, most DEGs associated with tumor-specific hyper-accessible regions were upregulated in tumors, while most DEGs associated with normal-specific hyper-accessible regions displayed downregulation (Fig. S3D, E). These findings indicate that the alterations in chromatin accessibility contribute to the differential expression of relevant genes in hepatocellular carcinoma.
DEGs associated with changes in chromatin accessibility are highly expressed in liver cancerTo further elucidate gene expression changes driven by chromatin accessibility alterations, we integrated the expression matrices of the four DAR groups and DEGs. Following correlation analysis and annotation of peaks to their nearest genes, we identified 22 DEGs significantly associated with changes in chromatin accessibility across the four groups (Table 2). Figure 2A depicts a correlation heatmap between chromatin accessibility and associated DEGs across the four sample groups. Notably, PRPF3 and PLIN2 emerged as consistently present across all four groups (Fig. 2B). The correlation analysis between the PRPF3 gene and its corresponding peak revealed a Pearson correlation coefficient of 0.57 (Fig. 2C). Visualization of the ATAC-seq data demonstrated that the PRPF3 transcription start site exhibited a higher peak intensity in tumor samples compared to normal tissues, signifying increased chromatin accessibility (Fig. 2D). Consistent with this finding, RNA-seq data revealed elevated PRPF3 expression in liver cancer (Fig. 2E). We further validated this observation by demonstrating elevated PRPF3 expression in liver cancer cell lines (Fig. 2F, G). Given the high PRPF3 expression in HCC, we investigated its potential as a prognostic biomarker. Analysis of TCGA data revealed a significant association between PRPF3 upregulation and poor patient prognosis (Fig. 2H). Additionally, immunohistochemical staining of four paired tumor and normal tissue samples demonstrated higher PRPF3 expression level in tumor tissues compared to normal tissues (Fig. 2I). These findings collectively suggest that PRPF3 may play a crucial role in HCC development and progression.
Table 2 Positively correlated DARs and DEGs in 4 sets of samplesFig. 2
Identification and expression analysis of differential genes correlated with chromatin accessibility changes positively. A The heatmap showing the differentially accessible regions (DARs) in chromatin and differentially expressed genes(DEGs) across the four pairs of samples. B The Venn diagram shows that PRPF3 and PLIN2 are the common DARs-associated DEGs across the four sample groups. C The correlation analysis between PRPF3 and its corresponding DARs. The Pearson’s correlation coefficient (R) and the corresponding P value are shown. D The panel showing the chromatin accessibility at the PRPF3 gene locus in 4 pairs of tumor and adjacent normal tissue samples, with red and blue boxes highlighting regions with differential accessibility between them. E PRPF3 expression level between HCC and normal samples in TCGA LIHC database. F, G The expression of PRPF3 in the HCC cell lines, including MHCC-97 h, Huh-7, HepG2, Jhh-7, and the normal liver cell line LO2 were investigated using qRT-PCR(F) and western blotting(G). H The Kaplan–Meier survival curves stratified by PRPF3 expression level. I Representative immunohistochemical staining of PRPF3 protein in 4 pairs of tumor and adjacent normal tissues (scale bar, 100 μm)
Knockdown of PRPF3 inhibits HCC cell proliferation, migration, and invasion in vitroTo investigate the functional role of PRPF3 in HCC, MHCC-97 h, and Huh7 cell lines were selected for further experimentation. Western blot and RT-qPCR analyses confirmed the efficacy of two independent siRNAs (siPRPF3-1 and siPRPF3-2) in downregulating PRPF3 expression at both the mRNA and protein levels (Fig. 3A, B). The CCK8 assay revealed a significant decrease in absorbance in the siPRPF3-1 and siPRPF3-2 groups compared to the siNC group at 24, 48, and 72 h, indicating reduced cell viability (Fig. 3C). Colony formation assays further demonstrated a decline in the colony formation ability of PRPF3-knockdown HCC cells (Fig. 3D). The EdU incorporation assay corroborated these findings, showing a reduction in the number of proliferating HCC cells following PRPF3 knockdown (Fig. 3E, F). Collectively, these results suggest that PRPF3 knockdown suppresses HCC cell proliferation. Next, Transwell assays were performed to assess migration. As shown in Fig. 3G and H, the number of cells migrating through the membrane was significantly reduced in the PRPF3 knockdown groups. Similarly, wound healing assays revealed a marked decrease in wound closure rate in the knockdown groups (Fig. 3I, J), demonstrating that PRPF3 knockdown also impairs the migration and invasion abilities of HCC cells.
Fig. 3
Knockdown of PRPF3 inhibits the proliferation, migration, and invasion of liver cancer cells in vivo. A, B The knockdown efficiency of PRPF3 was confirmed in the MHCC-97 h and Huh7 cell lines using qRT-PCR(A) and western blotting(B). C CCK8 analysis of cell viability in MHCC-97 h and Huh7 cells transfected with siNC, siPRPF3-1, or siPRPF3-2. D The colony formation images of MHCC-97 h and Huh7 cells transfected with siNC, siPRPF3-1, or siPRPF3-2 on day 14. E, F EdU proliferation assay in MHCC-97 h and Huh7 cells transfected with siNC, siPRPF3-1, or siPRPF3-2. G, H Transwell assay in MHCC-97 h and Huh7 cells transfected with siNC, siPRPF3-1, or siPRPF3-2. I, J Wound healing assay in MHCC-97 h and Huh7 cells transfected with siNC, siPRPF3-1, or siPRPF3-2
PRPF3 enhances HCC cell proliferation in vitro and in vivo and promotes migration and invasion in vitroHaving established that PRPF3 knockdown suppressed proliferation, migration, and invasion of HCC cells, we investigated whether its overexpression could conversely promote these processes. Stable PRPF3-overexpressing cell lines were established in MHCC-97 h and Huh7 cells via lentiviral transfection (Fig. S4A, B). CCK8, colony formation, and EdU incorporation assays consistently demonstrated that PRPF3 overexpression enhanced the proliferative capacity of HCC cells (Fig. S4C–E). Similarly, Transwell and wound healing assays revealed that PRPF3 overexpression promoted the migration and invasion abilities of HCC cells (Fig. S4F, G).
To assess the in vivo effects of PRPF3 on tumor progression, stable PRPF3-overexpressing Huh7 cells were subcutaneously injected into the right flank of nude mice. A control group received cells transfected with the empty pWPXL vector. Tumor volume and weight were significantly lower in the control group compared to the PRPF3-overexpressing group (Fig. S4H). These findings collectively demonstrate that PRPF3 can promote HCC cell proliferation both in vitro and in vivo.
The TF ZNF93 regulates the expression of PRPF3To identify potential upstream transcription factors regulating PRPF3 expression, we employed MEME FIMO to scan the DARs for transcription factor binding motifs. Given that transcription factors typically exert their regulatory effects by binding to gene promoter regions, we subsequently screened for factors with binding sites located within 2 kb upstream to 100 bp downstream of the PRPF3 transcription start site. Table 3 summarizes the top five transcription factors identified from each set of scanning results that satisfied these criteria. Notably, ZNF93 emerged as a potential transcription factor binding to the PRPF3 promoter region in all four DAR sets, exhibiting consistently high scores. To preliminarily validate the ZNF93-PRPF3 interaction, we utilized the Cistrome Data Browser database. The ChIP-seq results revealed a distinct enrichment peak for ZNF93 at the PRPF3 promoter region (Fig. 4A), suggesting potential binding of ZNF93 to the PRPF3 promoter, which displays increased chromatin accessibility in tumor tissues (Fig. 4B). Furthermore, western blot, RT-qPCR, and immunohistochemistry analyses demonstrated that ZNF93 expression is elevated in HCC cells and tissues (Fig. 4C–E). To functionally validate the regulatory role of ZNF93 on PRPF3 expression, we assessed PRPF3 levels in HCC cells with ZNF93 knockdown or overexpression. As expected, a positive correlation between PRPF3 and ZNF93 expression was observed (Fig. 4F–I). These findings collectively suggest that ZNF93 acts as an upstream transcription factor directly targeting and promoting PRPF3 expression.
Table 3 Transcription factors with binding sites located within the PRPF3 promoter regions in 4 sets of DARsFig. 4
ZNF93 positively regulates the expression of PRPF3 and is highly expressed in HCC. A The ChIP-seq data of ZNF93, the red boxes show a significant binding peak in the promoter region of PRPF3. B The panel shows the chromatin accessibility at the ZNF93 gene locus in 4 pairs of tumor and adjacent normal tissue samples, with red and blue boxes highlighting regions with differential accessibility. C, D The expression of ZNF93 in the HCC cell lines MHCC-97 h, Huh-7, HepG2, Jhh-7, and the normal liver cell line LO2 were investigated through qRT-PCR(C) and western blotting(D). E Representative immunohistochemical staining of ZNF93 protein in 4 pairs of tumor and adjacent normal tissues (scale bar, 100 μm). F, G The expression of PRPF3 was decreased in HCC cells after knocking down ZNF93. H, I The expression of PRPF3 was increased in HCC cells with ZNF93 overexpression
Knockdown of ZNF93 inhibits HCC cell proliferation, migration, and invasion in vitroTo investigate the functional role of ZNF93 in HCC cells, MHCC-97 h, and Huh7 cell lines were employed. Two independent siRNAs (siZNF93-1 and siZNF93-2) efficiently downregulated ZNF93 expression (Fig. 5A, B). Subsequent CCK8, colony formation, and EdU incorporation assays consistently demonstrated that ZNF93 silencing inhibited HCC cell proliferation (Fig. 5C–F). Similarly, Transwell and wound healing experiments revealed that ZNF93 knockdown suppressed both migration and invasion abilities of HCC cells (Fig. 5G–J).
Fig. 5
Knockdown of ZNF93 inhibits the proliferation, migration, and invasion of liver cancer cells in vivo. A, B The knockdown efficiency of ZNF93 was confirmed in the MHCC-97 h and Huh7 cell lines using qRT-PCR(A) and western blotting(B). C CCK8 analysis of cell viability in MHCC-97 h and Huh7 cells transfected with siNC, siZNF93-1, or siZNF93-2. D The colony formation ability of MHCC-97 h and Huh7 cells transfected with siNC, siZNF93-1, or siZNF93-2. E, F EdU assay in MHCC-97 h and Huh7 cells transfected with siNC, siZNF93-1, or siZNF93-2. G, H Transwell assay in MHCC-97 h and Huh7 cells transfected with siNC, siZNF93-1, or siZNF93-2. I, J Wound healing assay in MHCC-97 h and Huh7 cells transfected with siNC, siZNF93-1, or siZNF93-2
ZNF93 promotes HCC cell proliferation in vivo and in vitro, and enhances migration and invasion in vitroTo further investigate the functional consequences of ZNF93 overexpression on HCC cells, we established stable ZNF93-overexpressing cell lines in MHCC-97 h and Huh7 cells using lentiviral infection (Fig. S5A, B). CCK8, colony formation, and EdU incorporation assays consistently revealed that ZNF93 overexpression promoted the proliferation of HCC cells (Fig. S5C–E). Similarly, Transwell and wound healing experiments demonstrated that ZNF93 overexpression enhanced the migratory and invasive abilities of HCC cells (Fig. S5F–G). To assess the in vivo effects of ZNF93 on tumor growth, we employed a xenograft tumor model in immunodeficient mice. The results revealed that both tumor volumes and weights were significantly higher in the ZNF93-overexpressing group compared to the control group (Fig. S5H). These findings collectively indicate that ZNF93 can promote HCC cell proliferation both in vitro and in vivo.
PRPF3 is a target oncogene of ZNF93 in HCC cellsTo functionally validate whether PRPF3 functions as a downstream target of ZNF93 and mediates its oncogenic effects, we performed rescue experiments. Stable ZNF93-overexpressing Huh7 cells were subjected to PRPF3 knockdown using siPRPF3-1 and siPRPF3-2 (Fig. 6A). CCK8 and EdU incorporation assays demonstrated that PRPF3 downregulation effectively abrogated the ZNF93-induced proliferative effect (Fig. 6B, C). Transwell and wound healing assays further corroborated these findings, revealing that siPRPF3 impedes ZNF93-mediated enhancement of proliferation and migration in HCC cells (Fig. 6D, E). Collectively, these experimental results suggest that PRPF3 likely functions as a functional effector of ZNF93, mediating its tumor-promoting effects.
Fig. 6
Knockdown of PRPF3 in HCC cells overexpressing ZNF93 can suppress the oncogenic effects of ZNF93. A The protein expression profile of ZNF93 and PRPF3 in Huh7 cells overexpressing ZNF93 and with PRPF3 knockdown (siPRPF3-1, -2). B CCK-8 assay, C EdU assay, D Wound healing assay, and E Transwell assay were used to detect the effects of PRPF3 knockdown on the proliferation, migration, and invasion abilities of Huh7 cells stably overexpressing ZNF93
Comments (0)