
Remember me







Neurological disorders impact hundreds of millions of people around the world (Huang Y. et al., 2023). Neurological disorders encompass a wide range of conditions that impact the central, peripheral, and autonomic nervous systems of the body. Neurological disorders are a leading cause of disability and death worldwide (Huang Y. et al., 2023). Conditions like Alzheimer’s, Parkinson’s, stroke, and traumatic brain injuries can significantly impair quality of life and lead to premature mortality. The global prevalence of neurological disorders is projected to increase in the coming decades (Huang Y. et al., 2023). This is driven by aging populations, especially in low- and middle-income countries where the burden is often highest. The economic costs associated with neurological diseases are staggering, estimated at over $800 billion per year globally (Huang Y. et al., 2023). This includes direct medical costs as well as indirect costs from lost productivity. The challenges facing patients with nervous system disorders include irreversible damage, cognitive impairment, and resistance to treatment (Huang Y. et al., 2023). Additionally, the limited knowledge of the molecular causes of these conditions and the lack of early diagnostic tests and sensitive tools to monitor treatment effectiveness have significantly impeded available interventions, resulting in very poor prognoses for those with nervous system diseases. Improving our understanding of the molecular processes driving the development and progression of nervous system diseases, as well as promoting research into early diagnostic methods and new treatment approaches, are absolutely critical to addressing the substantial burden posed by these devastating neurological disorders.
Despite extensive research efforts, the limited engraftment and survival of transplanted stem cells continue to be significant challenges that must be overcome in order to improve the clinical efficacy of stem cell-based treatments for neurological disorders (Hoang et al., 2022; Temple, 2023; Zhang and Cheng, 2023). Recent research has shown that stem cells primarily exert their therapeutic effects on neurological diseases through the release of extracellular vesicles known as exosomes (Zhang and Cheng, 2023; Fayazi et al., 2021). These exosomes are rich in bioactive molecules, such as proteins, lipids, and ncRNAs, which collectively contribute to intercellular signaling and modulate various physiological processes. By enhancing cell survival, reducing inflammation, and promoting tissue repair, exosomes play a critical role in the regenerative potential of stem cells, offering promising avenues for the treatment of neurological disorders (Tan et al., 2024). Although the proportion of ncRNAs in Exo is relatively low, research shows that these ncRNAs, especially those present in stem cell-derived Exo, contribute significantly to the treatment of nervous system diseases by neuroprotection (Zhang L. et al., 2024),anti-apoptosis (Cheng et al., 2024), neuroplasticity and regeneration (Zhong et al., 2023), immunomodulation and anti-inflammation (Shan et al., 2023), epigenetic regulation and intercellular communication (Dragomir et al., 2018). For example, exosomal miR-146a-5p from umbilical cord-derived MSCs can suppress the activation of microglia and astrocytes, reducing neuroinflammation in models of ischemic stroke (Zhang Z. et al., 2021). Exosomal lncRNA MALAT1 from adipose-derived stem cells can stimulate the differentiation of neural stem/progenitor cells and enhance neurogenesis in the injured brain (El Bassit et al., 2017).
Adult-derived stem cells are more readily available, simpler to acquire, and do not present the same ethical dilemmas as stem cells derived from embryonic sources (Zakrzewski et al., 2019). Developing therapeutic approaches that utilize adult stem cell-derived exosomes and their non-coding RNAs is of utmost importance, as this holds great promise for treating a diverse range of debilitating neurological diseases, which pose a significant global health challenge. This review systematically explores the research progress and the underlying mechanisms by which diverse adult stem cell-derived exosomal ncRNAs can be leveraged for the treatment of nervous system diseases.
2 Stem cellsStem cells are a group of unspecialized cells that have the remarkable capacity to continually divide and renew themselves, as well as the potential to develop into various functional cell types that make up different tissues and organs (Zakrzewski et al., 2019). Stem cells are primarily categorized based on their ability to differentiate into different cell types, the source or origin from which they are derived, and their developmental progression or lineage (Zakrzewski et al., 2019). Stem cells can be classified according to their degree of potency, or ability to differentiate into different cell types (Zakrzewski et al., 2019). They range from totipotent cells that can give rise to all cell types, to pluripotent cells that can form many but not all cell types, to multipotent, oligopotent and unipotent cells that can form more limited sets of specialized cell type (Zakrzewski et al., 2019). Stem cells are found in both embryonic and adult tissues. The most prominent examples of pluripotent stem cells are embryonic stem cells (ESCs) (Calabrese, 2022) and induced pluripotent stem cells (iPSCs) (Aboul-Soud et al., 2021). On the other hand, adult tissues contain multipotent stem cells such as hematopoietic stem cells (HSCs) (Carroll and St Clair, 2018), mesenchymal stem cells (MSCs) (Samsonraj et al., 2017), neural stem cells (NSCs) (Urbán et al., 2019), and endothelial stem/progenitor cells (EPCs) (Chong et al., 2016). The various subtypes of stem cells, including embryonic, induced pluripotent, and adult multipotent stem cells, have been extensively studied and trialed as potential treatments for a wide range of human diseases. Exosomes are considered miniature versions or representations of their parent cells, in part because exosomes from a particular cell type contain cell-specific or unique sets of biomolecules. Additionally, stem cells have been found to function in a paracrine manner, meaning they act through the soluble factors they secrete, including exosomes, rather than solely through direct cell-cell interactions. In essence, the exosomes secreted by stem cells (SC-Exo) inherit similar therapeutic benefits as their parental stem cells, such as anti-inflammatory effects, immunomodulatory properties, and the ability to promote tissue regeneration (Tan et al., 2024). Stem cells have distinct ncRNA expression profiles that influence their fate and differentiation pathways (Marson et al., 2008). These unique expression profiles can be harnessed to guide the terminal differentiation of somatic cells from stem cells, providing a powerful strategy for regenerative medicine (Yu et al., 2023). By leveraging specific ncRNAs associated with particular cell types, researchers can effectively direct stem cells to develop into specialized cells. This approach holds significant promise for treating a variety of diseases, including muscle diseases, cardiovascular diseases, and neurological diseases. Stem cell-derived exosomes can share some common ncRNA profiles, but they also exhibit significant variability depending on the type of stem cell and the specific conditions under which they are derived (Tan et al., 2024). Many types of stem cells (e.g., embryonic stem cells, mesenchymal stem cells) may express certain core ncRNAs, such as specific miRNAs that are involved in regulating stemness, proliferation, and differentiation (Tan et al., 2024). The differences in ncRNAs among exosomes from various stem cell types can lead to distinct biological effects. For example, exosomes from mesenchymal stem cells might promote tissue repair, while those from neural stem cells might enhance neuroprotection (Tan et al., 2024). The local microenvironment and the specific stimuli (e.g., hypoxia, inflammation) can influence the composition of ncRNAs in exosomes, leading to differential effects on recipient cells. Each stem cell type has its unique transcriptional and post-transcriptional regulation mechanisms, leading to different ncRNA profiles. The presence of specific growth factors, cytokines, and other signaling molecules can alter ncRNA expression in stem cells, thus affecting the composition of their derived exosomes. The differentiation state of the stem cells (e.g., undifferentiated vs. differentiated) can also influence the ncRNA profiles, as the cells adapt their exosomal content to their functional roles. Epigenetic factors can play a significant role in regulating the expression of ncRNAs, contributing to the variability seen in exosome content among different stem cell types. However, using ncRNAs from exosomes derived from different stem cells has advantages when used as agents for neurological disorders (Pishavar et al., 2022). One advantage of exosomes derived from stem cells is that they can effectively pass through the BBB, enabling them to transmit ncRNAs to the nervous system. Moreover, by encapsulating the ncRNAs within exosomes, they become more stable and protected from degradation. Furthermore, exosomes derived from stem cells have features that can help in the regeneration of tissues, offer protection to the brain, and influence the microenvironment affected by diseases in the brain (Pishavar et al., 2022). This review focuses on understanding the role of ncRNAs obtained from exosomes derived from different stem cells in the treatment of various neurological disorders. The article emphasizes the importance of conducting research and investigation into these ncRNAs. Such exploration holds the potential for developing treatment strategies focusing on improving the healing process and minimizing complications in individuals with neurological disorders. Researchers are currently exploring the applications of stem cell-derived exosomes in neurological diseases, skin injuries, gastrointestinal diseases, and cancer (Figure 1). To better understand the functionality of these stem cell-derived exosomal ncRNAs-based therapies, further research is needed to develop improved techniques for the production and purification of the ncRNAs, as well as to establish robust protocols for their therapeutic delivery.
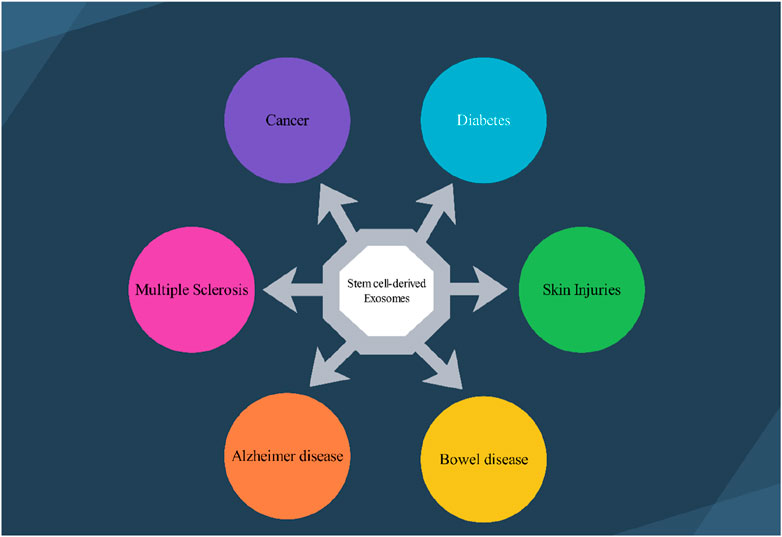
Figure 1. Several stem cell–derived exosomes therapeutic application in various disease. 1.1) Exosomes deliver neuroprotective factors, facilitating neuronal survival and regeneration in conditions like Alzheimer’s and Multiple Sclerosis’s disease. 1.2) Stem cell-derived exosomes can modulate the tumor microenvironment, potentially inhibiting tumor growth and enhancing the efficacy of chemotherapeutics. 1.3) Stem cell-derived exosomes are shown to improve insulin sensitivity and promote pancreatic beta-cell regeneration. 1.4) Stem cell-derived exosomes facilitate wound healing by promoting skin cell proliferation, angiogenesis, and reducing inflammation, thus enhancing the repair of damaged skin. 1.5) Exosomes play a role in modulating inflammation and promoting mucosal healing in conditions such as inflammatory bowel disease (IBD), aiding in the restoration of gut integrity.
3 Exosomes biogenesisExosomes originating from stem cells have shown promise in the treatment of various conditions (Zhang Q. et al., 2024; Huang Y. et al., 2023; Hoang et al., 2022). Exosomes are a specific subset of the broader category of extracellular vesicles (EV), distinguished by their smaller size range of 30–100 nm in diameter (Temple, 2023). The EV family encompasses several different types of membrane-bound structures, each with their own unique properties and functions. Exosomes carry a diverse cargo of biomolecules, including proteins, lipids, and nucleic acids, that are reflective of the parent cell that produced the exosome. This biomolecular content within exosomes can function as distinctive “markers” or “signatures” that provide information about the cell of origin. Exosomes commonly contain protein markers such as tetraspanins (CD9, CD63, CD81) and membrane transport/fusion proteins like Rab GTPases and SNAREs (Figure 2) (Temple, 2023). Heat shock proteins are also frequently detected as exosomal markers (Temple, 2023). The specific molecular composition of exosomes reflects the unique makeup of the parent cell, providing valuable information about the state and origin of the exosomes. Exosomes are formed and secreted through a multi-step process. Recent evidence indicates that exosomes are generated not only through the endosomal pathway, but also via a process of membrane invagination. The exosome formation process begins with the internalization of various extracellular materials, including proteins, lipids, metabolites, and cell membrane components, into the cell via endocytosis (Figure 3) (Zhang and Cheng, 2023). The internalization occurs through the invagination, or inward folding, of the cell membrane, creating primary endocytic vesicles that enclose the extracellular materials. The primary endocytic vesicles then fuse together to form early endosomes (EEs) within the cell. These early endosomes subsequently undergo a maturation process, transforming and developing into late endosomes (LEs). As the late endosomes mature, their internal membrane begins forming multiple invaginations or inward folds within the lumen of the organelle. The selective membrane invaginations within the late endosomes encapsulate and sort specific proteins, lipids, nucleic acids, and other cellular components, packaging them into intraluminal vesicles (ILVs) inside the organelle. The late endosomes containing the ILVs then further develop and mature into multivesicular bodies (MVBs). Specific MVBs are directed towards the cell’s outer membrane by intracellular transport mechanisms. These MVBs then merge with the plasma membrane, releasing their internal vesicle contents into the extracellular space. These released vesicles are now classified as exosomes, which can be absorbed by nearby cells or continue to circulate outside the cell. Some MVBs are transported to and merge with the cell’s lysosomes. This enables the contents of the MVBs, including any unwanted or damaged materials, to be broken down and cleared from the cell. The process is regulated by a coordinated effort between key proteins, including RAB GTPases, tethering factors, SNARE proteins, and cytoskeletal proteins. These proteins work together to ensure the MVBs are properly transported, docked, and fused with the cell’s outer membrane, facilitating the release of exosomes (Zhang and Cheng, 2023). In addition to the regulatory proteins, important environmental factors within the cell - including calcium levels, pH, and oxygen concentrations - also significantly impact the production and release of MVBs and exosomes. Conditions such as high calcium, low pH, and low oxygen levels can all enhance and regulate the formation and release of these extracellular vesicles (Fayazi et al., 2021). The interplay between the specialized proteins involved and the cellular environmental factors is critical for ensuring the proper and directed secretion of exosomes.
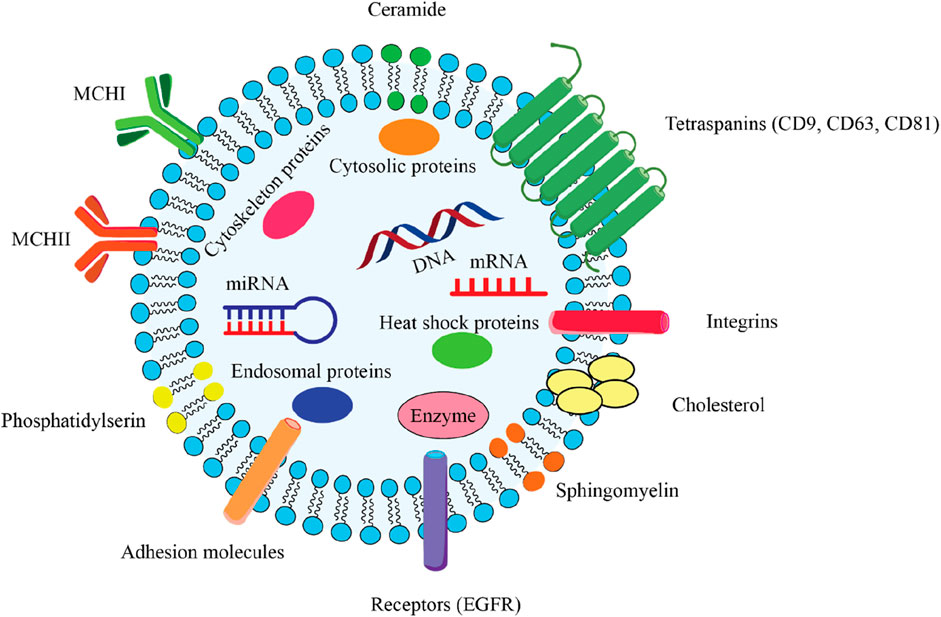
Figure 2. Comprehensive overview of the key structural components of an exosome, which is a type of extracellular vesicle involved in intercellular communication. 2.1) Lipid bilayer membrane: The outer structure of the exosome, composed of a phospholipid bilayer that encases the internal cargo and maintains the integrity of the vesicle. 2.2) Encapsulated Cargo: mRNA and miRNA play critical roles in gene regulation and protein synthesis. DNA that can be transferred between cells. Cytosolic proteins include various proteins that are important for cellular function and signaling. Endosomal proteins derived from the endosomal pathway, essential for exosome biogenesis. Enzymes catalysts biochemical reactions that may influence recipient cells. 2.3) Surface Markers: Tetraspanins (CD9, CD63, CD81) facilitate exosome formation and play a role in cell recognition and adhesion. Integrins mediate interactions with the extracellular matrix and other cells. Adhesion molecules that assist in the binding of exosomes to target cells. Receptor (EGFR) can mediate cellular responses upon binding with ligands. 2.4) Lipid Components: Ceramide is a sphingolipid that contributes to membrane stability and signaling. Cholesterol is a critical component for maintaining membrane fluidity and structure. Phosphatidylserine is a phospholipid that is involved in cell signaling and recognition. Sphingomyelin is a type of sphingolipid that plays a role in membrane structure and function. 2.5) Heat Shock Proteins: These proteins are involved in protecting cells from stress and facilitating protein folding.
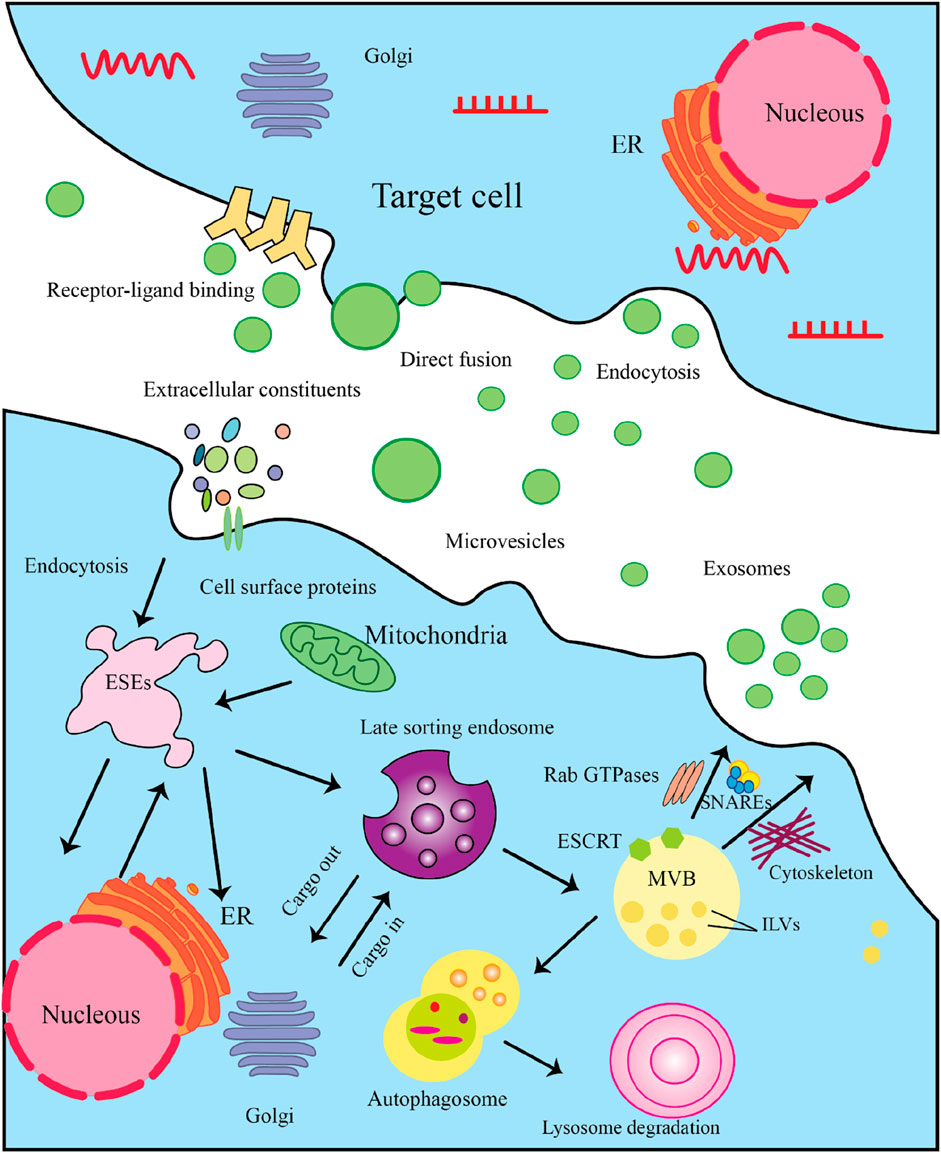
Figure 3. The Multistep Process of Exosome Biogenesis and Secretion. Here’s a detailed explanation of the steps involved. 3.1) Cargo Synthesis and Sorting: The process begins in the nucleus, where mRNAs and proteins are synthesized. These molecules are then translated in the endoplasmic reticulum (ER). After synthesis, proteins are transported to the Golgi complex for post-translational modifications and sorting. 3.2) Formation of Exosome-Secretory Endosomes (ESEs): Cells can internalize extracellular constituents through endocytosis. This process allows the uptake of cellular materials, including proteins and lipids, which may be incorporated into exosomes. Some membrane protrusions can shed from the cell surface, forming microvesicles that may also participate in intercellular communication. 3.3) Late Sorting Endosome: Internalized materials are directed to late endosomes, which serve as sorting hubs for cargo destined for exosomes. Here, the endosomal membrane invaginates to form intraluminal vesicles (ILVs). The Endosomal Sorting Complex Required for Transport (ESCRT) is crucial for the budding of ILVs within late endosomes, facilitating the encapsulation of proteins and nucleic acids into these vesicles. 3.4) MVB Formation: As ILVs accumulate within late endosomes, they transform into multivesicular bodies (MVBs). MVBs can either fuse with lysosomes for degradation or with the plasma membrane for exosome release. 3.5) Exosome Secretion: MVBs that are destined to release exosomes will fuse with the plasma membrane, allowing ILVs to be expelled as exosomes into the extracellular space. In some cases, exosomes may also enter target cells by directly fusing with the cell membrane, releasing their cargo. 3.5) Target Cell Interaction: Once in the extracellular environment, exosomes can interact with target cells through specific receptor-ligand binding, which initiates signaling pathways. Target cells can uptake exosomes through endocytosis, allowing the internalization of their bioactive molecules.
4 Exosome-derived noncoding RNAsExosomes contain a wide range of ncRNA types, including miRNAs, lncRNAs, circRNAs, piRNAs, and siRNAs. As the discovery and study of non-coding RNAs (ncRNAs) in exosomes has advanced, many innovative functions and applications have emerged. These range from novel mechanisms of cell-to-cell communication to the use of exosomal ncRNAs as promising disease biomarkers. Furthermore, exosomal ncRNAs have shown new therapeutic applications, particularly in the context of neurological diseases. The incorporation of specific ncRNAs into exosomes is a highly regulated process, involving various cellular mechanisms and pathways. For instance, the endosomal sorting complex required for transport (ESCRT) proteins play a crucial role in the incorporation of miRNAs into intraluminal vesicles (ILVs), which are then released as exosomes (Henne et al., 2011; Frankel and Audhya, 2018). The ESCRT machinery is involved in the budding and scission of the ILVs, providing a mechanism for the selective packaging of miRNAs into these vesicles (Frankel and Audhya, 2018). Also, the neutral sphingomyelinase 2 (nSMase2) enzyme has been implicated in the sorting of miRNAs into exosomes (Frankel and Audhya, 2018). nSMase2 catalyzes the hydrolysis of sphingomyelin to ceramide, which can promote the formation of lipid-enriched microdomains in the endosomal membrane. These ceramide-enriched microdomains may then facilitate the recruitment and packaging of miRNAs into the ILVs. These two pathways highlight the complex and regulated nature of the processes involved in the loading of specific ncRNA species into these extracellular vesicles. The detailed mechanisms underlying the selective packaging of various ncRNA species into exosomes require further exploration and research.
4.1 microRNAsmiRNAs are short, ncRNA molecules that do not encode proteins, typically ranging from 19 to 25 nucleotides in length, and play regulatory roles in eukaryotic organisms (Ranganathan and Sivasankar, 2014). In the nucleus, the genes encoding miRNAs are transcribed into long initial transcripts (pri-miRNAs), which are then processed by the Drosha and DGCR8 proteins to produce a stem-loop structure called pre-miRNA, approximately 70 nucleotides long (Ranganathan and Sivasankar, 2014; Michlewski and Cáceres, 2019). This pre-miRNA is further cleaved by the Dicer enzyme in the cytoplasm to generate the mature miRNA. The mature miRNA is then exported from the nucleus and, with the help of a helicase, can assemble into the RNA-induced silencing complex (RISC) (Iwakawa and Tomari, 2022). Within the RISC complex, the miRNA can regulate gene expression by base pairing with complementary sequences on target mRNAs, leading to mRNA degradation or translational repression.
4.2 Long non-coding RNAslncRNAs are a class of ncRNAs that are longer than 200 nucleotides, with some exceeding thousands of nucleotides in length (Han and Chen, 2024). Unlike miRNAs, lncRNAs can adopt complex secondary and tertiary structures, which are important for their diverse functional roles. The subcellular localization of lncRNAs is an important factor that determines their diverse functional roles in eukaryotic cells. In the cell nucleus, lncRNAs can participate in diverse regulatory functions, including transcriptional regulation, organization of chromatin structure, and regulation of RNA splicing (Han and Chen, 2024). In the cytoplasm, such as in exosomes, they can serve different roles, including post-transcriptional regulation, sequestration of microRNAs, and various signaling functions (Han and Chen, 2024). Concurrently, the expanding knowledge about exosomal long non-coding RNAs has driven increased focus on understanding their roles in biology. This has steadily revealed that exosomal lncRNAs play a significant part in the treatment of many nervous system diseases.
4.3 CircRNAsCircRNAs are a form of non-coding RNA that are produced through a reverse splicing mechanism and reside within the cell nucleus (Feng et al., 2023). Certain circRNAs are derived from the non-coding segments of genes, while others include at least one protein-coding region and are predominantly located in the cell’s cytoplasm (Feng et al., 2023). CircRNAs are abundantly present in tissues and cell types, including the brain and have been conserved across different species throughout evolution. Recent research has revealed that exosomal circular RNAs can serve multiple functions, including acting as regulating splicing and transcription, inhibiting proteins, and miRNA sponges. They play roles in biological processes and are implicated in several disease conditions. In the field of neurobiology, specifically circRNAs, there is an increasing body of reports suggesting their role in the progression of disorders such as Parkinson’s disease, schizophrenia, Alzheimer’s disease, and epilepsy. Researchers suggests that circRNAs are beneficial in identifying and addressing these conditions as both biomarkers (Cheng et al., 2024) and treatment targets (Cheng et al., 2024; El Bassit et al., 2017).
4.4 piRNAspiRNAs are a class of small non-coding RNA molecules that interact with PIWI proteins (Cheng et al., 2024). piRNAs guide PIWI proteins to complementary sequences, leading to the degradation or transcriptional silencing of target RNAs (Wu et al., 2023). The biogenesis of piRNAs involves a distinct pathway from miRNA or siRNA production and are thought to be important for germ cell development, stem cell maintenance, and epigenetic regulation (Wu et al., 2023). In contrast to microRNAs, piRNAs have the ability to silence genes. They can participate in diverse processes, such as transcriptional silencing or activation, transposon silencing, post-transcriptional regulation, and other modifications (Wu et al., 2023). This highlights the unique characteristics of piRNAs compared to other types of non-coding RNAs. piRNAs appear to be important players in maintaining neuronal homeostasis, supporting brain development, and potentially contributing to cognitive processes and neurological health (Zhong et al., 2023).
5 NcRNAs from stem cell–derived exosomes and their potential mechanisms in neurological disordersNcRNAs obtained from stem cell-derived exosomes can exert therapeutic effects against neurological disorders via different mechanisms. Here are some possible mechanisms by which these ncRNAs from stem cell–derived exosome regulate biological functions and promote neurological outcomes (Figure 4).
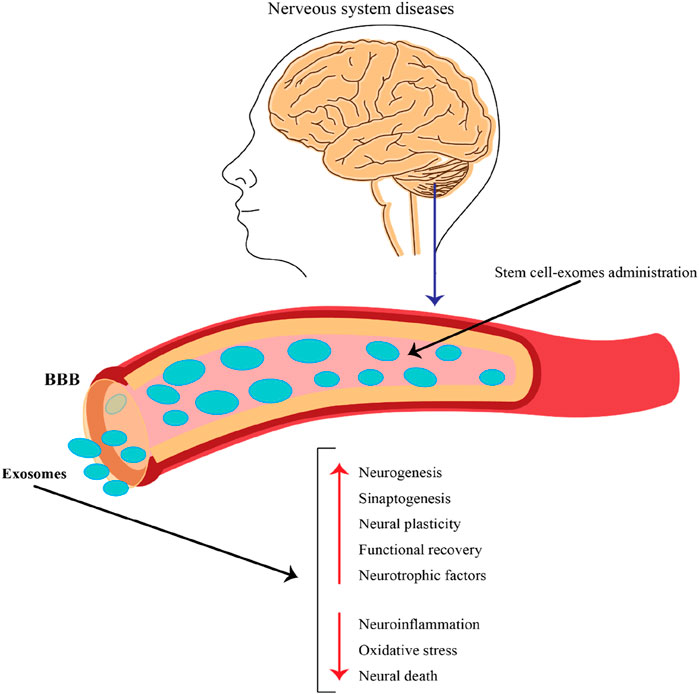
Figure 4. The role of exosomes released by stem cells in the context of nervous system diseases and their potential therapeutic applications. This figure emphasizes the potential of stem cell-derived exosomes as a therapeutic strategy for treating nervous system diseases. By crossing the BBB and delivering beneficial cargo, these exosomes can promote neural repair, enhance recovery, and combat harmful processes in the brain, making them a promising avenue for future research and clinical applications.
5.1 Modulation gene expression patterns associated with key neural processesStem cell-derived exosome ncRNAs can modulate the expression of specific genes by binding to mRNA molecules (Tian et al., 2022; Bai et al., 2022; Xie et al., 2023) and altering their stability or translation. Through molecular interactions, ncRNAs can influence the expression of genes associated with processes like synaptic plasticity (Pan et al., 2020), neuroprotection (Zhuang et al., 2023; Wang et al., 2021), neuroinflammation (Zhang Z. et al., 2021; Zhang Y. et al., 2021), neurogenesis (Zhang Y. et al., 2021; Pan et al., 2021), and other significant biological processes.
5.1.1 NeuroinflammationNeurological disorders often exhibit a characteristic called neuroinflammation. This phenomenon occurs when immune cells become active and liberate substances that prompt inflammation. Stem cell-derived exosomes can transport ncRNAs, including miRNAs and lncRNAs, which can specifically target genes associated with immune responses and inflammation (Zhang Z. et al., 2021; Zhang Y. et al., 2021). By controlling the expression of these genes, stem cell-derived exosomal ncRNAs can effectively modify the response in the nervous system and reduce neuroinflammation associated with neurological disorders (Zhang Z. et al., 2021; Zhang Y. et al., 2021).
5.1.2 NeuroprotectionExperimental studies confirmed that exosomes derived from stem cells, along with their ncRNAs content, can support the nervous system in various neurological disorders. These exosomes can transport ncRNAs, which target genes involved in cell survival (El Bassit et al., 2017; Pan et al., 2020; Liu J. et al., 2022), apoptosis, and response to oxidative stress. By modifying the expression of these genes, the ncRNAs carried by exosomes can protect neurons’survival, decrease cell death rates, and increase the resilience of neurons against insults. This effects finally helps to safeguard the system during disease conditions (Chang et al., 2021; Luo et al., 2022).
5.1.3 Synaptic plasticityLearning, memory, and general brain function are all dependent on synaptic plasticity, or the synapses’ ability to change in strength and connectivity. Exosomes derived from stem cells can convey ncRNAs that regulate the expression of genes implicated in synaptic plasticity (Pan et al., 2020). These genes encode neurotransmitter receptors, synaptic proteins, and signaling molecules. Exosomal ncRNAs may improve synaptic plasticity (Pan et al., 2020) and neuronal connectivity in neurological disorders with synaptic dysfunction by regulating the expression of these genes.
5.1.4 Neurogenesis and differentiationNeurogenesis is crucial for brain development, learning, and memory formation. Stem cells, which have the ability to differentiate into various cell types, play a crucial role in neurogenesis. NcRNAs containing exosomes derived from stem cells can control the expression of genes that play a role in these processes, encouraging the growth of neurons. The development of spines, the formation of synapses, and the modification of existing synaptic connections. By enhancing these processes, stem cell-derived exosomal ncRNAs can contribute to neuronal regeneration (Zhang L. et al., 2024; Tan et al., 2024; Pishavar et al., 2022; Henne et al., 2011), functional recovery (Zhang L. et al., 2024; Sun and Xu, 2023), and cognitive improvement (Zhang L. et al., 2024; Tan et al., 2024; Huang et al., 2021) in neurological disorders. Stem cell-derived exosomes can transport ncRNAs that target genes involved in NSCs fate determination (Zhang L. et al., 2024; Tan et al., 2024). Adult mammalian neural stem cells possess distinctive characteristics that set them apart from other stem cell types. These key properties include their ability to differentiate into various neural cell lineages, their capacity for self-renewal to maintain the stem cell pool, and their ability to enter a dormant state (Dong et al., 2022). NcRNAs containing exosomes derived from stem cells can stimulate neurogenesis through several key mechanisms. Exosomal ncRNAs can target and modulate the expression of key transcription factors involved in neural lineage specification and neurogenesis, such as SOX2, NEUROG2, ASCL1, and OLIG2. For example, exosomal miR-124 from NSCs can target the 3′ UTR of the SCP1 gene, which in turn upregulates the expression of the neurogenic transcription factor NEUROG2, promoting neuronal differentiation (Zhang et al., 2023). Exosomal ncRNAs can regulate signaling pathways that are crucial for NSCs fate determination, proliferation, and neuronal differentiation, such as the Notch, Wnt, and Sonic Hedgehog pathways. Exosomal ncRNAs can influence epigenetic modifications, such as DNA methylation and histone acetylation, which can alter the expression of genes involved in neurogenesis. Exosomal lncRNA MALAT1 from NSCs has been reported to interact with the polycomb repressive complex 2 (PRC2), leading to the epigenetic silencing of genes that inhibit neuronal differentiation (Zhao C. et al., 2022). Exosomal ncRNAs can target genes and pathways involved in neuronal maturation, dendritic arborization, and synaptic formation and function. Exosomal miR-132 from MSCs has been shown to enhance dendritic spine formation and synaptic plasticity, contributing to the functional integration of newborn neurons (Ma et al., 2022). Specific exosomal ncRNAs can modulate the inflammatory response in the central nervous system, which can influence the neurogenic niche and support the survival and integration of newborn neurons. Exosomal lncRNA NEAT1 from NSCs has been implicated in the regulation of neuroinflammation, potentially promoting a more favorable environment for neurogenesis (Zhang Q. et al., 2024). Exosomal ncRNAs can be transferred to other cell types in the neurogenic niche, such as astrocytes, oligodendrocytes, and endothelial cells, influencing their functions and indirectly supporting neurogenesis. For instance, exosomal miR-21 from NSCs can be taken up by astrocytes, leading to the upregulation of neurotrophic factors that support neuronal survival and differentiation (Zhong et al., 2023).
5.1.5 Blood-brain barrier integrityBBB plays a critical role in modulating the passage of molecules and cells into the brain. Compromised BBB leads to the development of neurological disorders. However, there is a potential solution using exosomes derived from stem cells. These exosomes convey special ncRNAs that can impact the expression of genes responsible for preserving BBB integrity and function. By modulating these gene expressions, exosomal ncRNAs can effectively support the BBB barrier against substances entering the brain (Wang et al., 2022; Hill, 2019).
5.2 Modulation of signaling pathwaysStem cell-derived exosomal ncRNAs can influence critical signaling pathways involved in the development of neurological disorders. As an example, miRNAs have the ability to change the function of signaling pathways, including the Wnt/β-catenin (Xu et al., 2019), AKT/ERK pathways (Zhuang et al., 2023; Jiang et al., 2021), and PI3K/Akt pathways (Pan et al., 2020; Wei et al., 2020). These pathways closely play critical roles in cellular processes such as synaptic plasticity, neurogenesis, and cell survival. By modulating these pathways, ncRNAs can affect cellular processes related to neurological disorders and facilitate neuroprotection and functional recovery. The modification of signaling pathways is a crucial component of cellular function and is frequently disrupted in neurological disorders. Stem cell-derived exosome ncRNAs have been identified to regulate signaling pathways in neurological disorders. The following sections provide an overview of how ncRNAs from different stem cell-derived exosomes affect signaling pathways and their implications for neurological disorders:
5.2.1 Wnt/β-catenin pathwayDisruption in the Wnt/β pathway may affect on brain development, synaptic plasticity, and the generation of neurons. Stem cell-derived exosomes contain ncRNAs that specifically target various elements of the Wnt/β-catenin pathway, such as β-catenin and Wnt ligands. These exosomal ncRNAs can regulate the expression or activity of these targets, consequently influencing the signaling of the Wnt/β-catenin pathway. Consequently, this modulation has implications for safeguarding the health of the human brain, the adapting our connections, and generating new brain cells (Xu et al., 2019).
5.2.2 PI3K/Akt/mTOR pathwayThe PI3K/Akt/mTOR pathway plays a critical role in promoting the survival of neurons, their growth, and the plasticity of synapses. Stem cell-derived exosomes can carry ncRNAs that target genes in this pathway, such as PI3K, Akt, and mTOR. By modulating the expression or activity of these genes, exosomal ncRNAs can modulate PI3K/Akt/mTOR signaling, influencing survival, dendritic growth, and synaptic connectivity in neurological disorders (Wei et al., 2020).
5.2.3 AKT/ERK pathwaysThe AKT/ERK pathway is essential for neuronal development, synaptic plasticity, and neuronal survival. Stem cell-derived exosomes can transport ncRNAs that target components of this pathway, including kinases and transcription factors (Zhuang et al., 2023; Jiang et al., 2021). Stem cell-derived exosomal ncRNAs can influence the expression or function of these elements, resulting in the modulation of AKT/ERK pathway signaling and its subsequent impact on function, synaptic plasticity, and neuroprotection (Zhuang et al., 2023; Wang et al., 2021).
5.2.4 NF-κB pathwayStem cell-derived exosomal ncRNAs can influence the expression or function of these elements, resulting in the modulation of NF-κB pathways signaling (Zhang Z. et al., 2021; Zhuang et al., 2023; Wang et al., 2021; Jiang and Zhang, 2021; Sun and Xu, 2023) and its subsequent impact on function, synaptic plasticity, and neuroprotection. Exosomes derived from stem cells can transport ncRNAs that specifically target genes involved in the NF-κB pathway, including NF-κB subunits,and upstream regulators (Wang et al., 2021; Jiang and Zhang, 2021). By modulating the expression or activity of these genes, stem cell-derived exosomal ncRNAs can influence NF-κB signaling and modulate neuroinflammation,and immune responses in neurological disorders (Jiang and Zhang, 2021; Sun and Xu, 2023; Fan et al., 2021).
The delivery of ncRNAs containing exosomes derived from stem cells to recipient cells provides a means to regulate the functioning of signaling pathways implicated in neurological disorders. By targeting the critical components of these pathways, ncRNAs containing exosomes derived from stem cells can influence various cellular processes, including neurodevelopment, neuroinflammation, synaptic plasticity (Pan et al., 2020), and neuroprotection. Additional investigations are required to reveal the ncRNAs implicated, their functions, and their potential for therapeutic influence on signaling pathways associated with neurological disorders.
5.3 Neuroinflammation modulationNumerous neurological disorders are influenced by neuroinflammation, which plays a crucial role in neurological development. Exosomes produced from stem cells are included in ncRNAs, which can target inflammatory pathways and control the production of pro- or anti-inflammatory proteins. By modulating neuroinflammatory responses, these ncRNAs can reduce inflammation (Zhang Z. et al., 2021; Zhang Y. et al., 2021), attenuate glial activation, and create a more favorable environment for neuronal survival (El Bassit et al., 2017; Pan et al., 2020; Liu J. et al., 2022) and regeneration (Zhang Y. et al., 2021; Pan et al., 2021; Huang et al., 2021; Patel et al., 2018). Below is a summary of the effect of ncRNAs derived from exosomes originating from stem cells on neuroinflammation and the implications for neurological disorders:
5.3.1 Regulation of inflammatory responsesExosomes originating from stem cells possess the capacity to transport specific types of ncRNAs, such as miRNAs and lncRNAs, which selectively target genes associated with immune responses and inflammation (Zhang Z. et al., 2021; Tian et al., 2022; Zhang Y. et al., 2021). These exosomal ncRNAs can regulate the levels of inflammatory substances in the body, such as cytokines (such as interleukins and tumor necrosis factor-alpha) and chemokine (Tian et al., 2022). They also affect the production of mediator-producing enzymes, such as cyclooxygenases and inducible nitric oxide synthases. By downregulating the expression of these pro-inflammatory molecules (Jiang and Zhang, 2021; Shao et al., 2020; Dong et al., 2022), exosomal ncRNAs can attenuate neuroinflammation in neurological disorders.
5.3.2 Regulation of glial activationGlial cells, such as microglia and astrocytes, have functions in processes related to inflammation in the brain. Stem cell-derived exosomes can carry ncRNAs that target genes involved in the activation and polarization of glial cells. For instance, specific exosomal miRNAs can control the activation of microglia, causing them to transition from an inflammatory (M1) state to an anti-inflammatory (M2) state (Chang et al., 2021; Shao et al., 2020; Dong et al., 2022; Yang et al., 2022; Zhao et al., 2020). By modulating the activation and polarization of glial cells (Zhang Z. et al., 2021; Xiaoying et al., 2020), the ncRNAs contained within stem cell-derived exosomes can regulate the immune responses and help reduce neuroinflammation.
5.3.3 Modulation of neuroinflammatory signaling pathwaysNeuroinflammation involves he activation of pathways such as the IRAK1/TRAF6 signaling pathway (Zhang Z. et al., 2021). Stem cell-derived exosomes can transport ncRNAs that target components of these pathways, thereby affecting their activity and downstream signaling. By modulating the activity of these signaling pathways, exosomal ncRNAs can influence the expression of pro-inflammatory genes and modulate neuroinflammatory responses (Zhang Z. et al., 2021).
5.3.4 Regulation of immune cell functionImmune cells, including microglia and infiltrating immune cells, play critical roles in neuroinflammation. Exosomes produced from stem cells can deliver ncRNAs to cells of the immune system, thereby modulating their function and behavior. For instance, exosomal ncRNAs can influence the phenotype and activation state of microglia (Zhang Z. et al., 2021; Xiaoying et al., 2020), and also regulate the recruitment and activation of peripheral immune cells. By modulating immune cell function, exosomal ncRNAs can influence neuroinflammatory processes in neurological disorders (Zhang Z. et al., 2021; Xiaoying et al., 2020).
5.4 Intercellular communication and paracrine effectsNcRNAs found in stem cell-produced exosomes can be absorbed by recipient cells, thereby changing the function of those cells. By transferring their content, these exosomes can regulate gene expression, signaling pathways, and various tasks within the recipient cells (Figure 5). This communication between cells can have effects and contribute to therapeutic benefits for nearby cells and tissues. Now let’s take a look at how ncRNAs from stem cell-derived exosomes impact communication and explore their potential implications for neurological disorders.
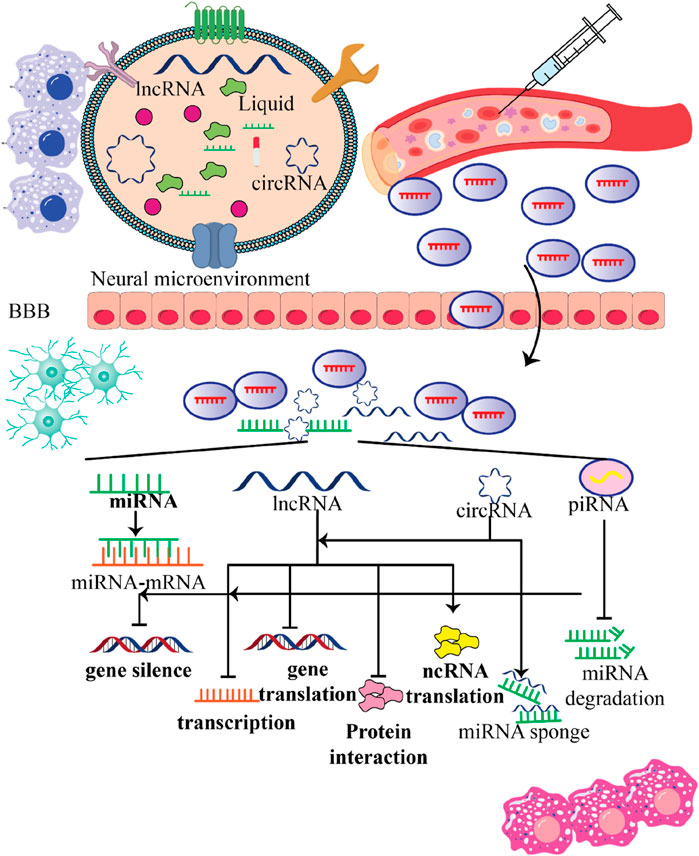
Figure 5. Various functions stem cell-produced exosomes contain ncRNAs within the recipient cells. This figure emphasizes the multifunctional roles of ncRNAs contained in stem cell-derived exosomes within the neural microenvironment. By crossing the blood-brain barrier (BBB) and influencing gene expression in recipient cells, these exosomes hold significant potential for therapeutic applications in treating nervous system disorders, highlighting their importance in innovative medical strategies.
5.4.1 Transfer of ncRNAsExosomes obtained from stem cells have the capability to transport various categories of ncRNAs, including miRNAs, lncRNAs, and circRNAs. The nervous system’s recipient cells, such as endothelial cells (Wang et al., 2022; Hill, 2019; Venkat et al., 2018), microglia (Chang et al., 2021; Shao et al., 2020; Dong et al., 2022; Yang et al., 2022; Zhao et al., 2020), neurons (Zhuang et al., 2023; Pan et al., 2021; Zhao Y. et al., 2022; Liu Y. et al., 2022), and astrocytes (Huang W. et al., 2023), can receive these ncRNAs from stem cells. These exosomal ncRNAs can influence gene expression and cellular function in the recipient cells that absorb the exosomes.
5.4.2 Regulation of gene expressionStem cell-derived exosomal ncRNAs can regulate gene expression in recipient cells through mechanisms. For instance, miRNAs have the ability to bind to mRNAs and cause translation inhibition or mRNA destruction. LncRNAs can change gene expression at both the transcriptional and non-transcriptional levels by interacting with proteins, RNA, or DNA. Through these actions that change gene expression patterns, exosomal ncRNAs have the potential to impact processes relevant to neurological disorders, including neuroinflammation, neurogenesis (Zhang Y. et al., 2021; Pan et al., 2021), synaptic plasticity, and neuronal survival (El Bassit et al., 2017; Pan et al., 2020; Liu J. et al., 2022).
5.4.3 Modulation of cellular phenotypesExtracellular ncRNAs can influence the characteristics of cells by controlling their development, growth, and overall functions. For instance, ncRNAs found in exosomes from stem cells can support the transformation of progenitor cells into neurons, improve survival rates (El Bassit et al., 2017; Pan et al., 2020; Liu J. et al., 2022), guide microglia toward an anti-inflammatory state (Chang et al., 2021; Shao et al., 2020; Dong et al., 2022; Yang et al., 2022; Zhao et al., 2020), or stimulate angiogenesis (Bai et al., 2022; Zhang Y. et al., 2021; Wang et al., 2022; Jiang et al., 2021). These actions play a role in facilitating tissue healing, protecting the system, and promoting functional restoration in neurological disorders.
5.4.4 Regulation of signaling pathwaysExosomes, which are membrane-bound vesicles, contain ncRNAs that can impact the behavior of recipient cells by affecting various cellular processes. One interesting example is how stem cell-derived exosomal miRNAs can interact with components of signaling pathways like Wnt/β (Xu et al., 2019), mTOR (Wei et al., 2020; Huang W. et al., 2023), NF-κB (Zhang Z. et al., 2021; Zhuang et al., 2023; Jiang and Zhang, 2021; Sun and Xu, 2023), NGF/TrkA (Wang et al., 2021), PI3K/Akt (Pan et al., 2020), or AKT/ERK (Jiang et al., 2021) pathways. This interaction can then influence responses related to synaptic plasticity and neuroinflammation. By modulating signaling pathways in the recipient cells, the stem cell-derived exosomal ncRNAs play a role in regulating various cellular functions. This modulation has important implications for understanding and potentially treating neurological disorders.
5.4.5 Neuroprotection and tissue repairExosomes produced from stem cells can convey ncRNAs that are essential for enhancing neuroprotection and expediting tissue restoration (Liu J. et al., 2022; Chang et al., 2021; Shao et al., 2020) in various neurological disorders. The presence of stem cell-derived exosomal ncRNAs enhances cell survival (El Bassit et al., 2017; Pan et al., 2020; Liu J. et al., 2022), reduces stress (Tian et al., 2022; Wang Q. S. et al., 2023), regulates inflammation, stimulates the formation of new blood vessels (Bai et al., 2022; Zhang Y. et al., 2021; Wang et al., 2022; Jiang et al., 2021), and facilitates tissue remodeling. These beneficial effects contribute to the preservation of function, restoration of damaged tissue, and overall promotion of recovery in individuals with disorders. The functional mechanisms of ncRNAs can vary depending on the specific type of ncRNAs they interact with and the particular neurological disorder being investigated. More research is necessary to understand the mechanisms through which ncRNAs from exosomes derived from stem cells confer therapeutic benefits. In addition, optimizing delivery methods for use requires exploration.
5.4.6 NcRNAs from stem cell-derived exosome and neurological disordersNcRNAs obtained from exosomes derived from stem cells have shown encouraging prospects concerning neurological disorders. These ncRNAs have the capability to regulate gene expression and manipulate cellular processes in the cells to which they are transferred, presenting potential therapeutic advantages. Because they can alter gene expression and neuroinflammation (Zhang Z. et al., 2021; Zhang Y. et al., 2021), ncRNAs produced from stem cell-derived exosomes have therapeutic potential in neurological diseases (Zhang Z. et al., 2021; Zhang Y. et al., 2021; Elballal et al., 2024), neurodegenerative diseases (Zhang Y. et al., 2021; Pan et al., 2021; Huang et al., 2021; Patel et al., 2018), and synaptic plasticity (Pan et al., 2020; Hu and Li, 2017). By targeting specific molecular pathways and cellular processes, these ncRNAs can exert neuroprotective effects (Xie et al., 2023; Pan et al., 2020; Zhuang et al., 2023; Luo et al., 2022; Losurdo et al., 2020), enhance functional recovery, and slow disease progression. To fully understand the mechanisms of action, optimize delivery methods, and ascertain the effectiveness and safety of stem cell-derived exosomes in human patients, more investigation and clinical trials are still needed. We present a few examples of neurological disorders associated with the involvement of ncRNAs derived from stem cell exosomes (Zhang Z. et al., 2021; El Bassit et al., 2017; Tian et al., 2022; Bai et al., 2022; Xie et al., 2023; Pan et al., 2020; Zhuang et al., 2023; Wang et al., 2021; Zhang Y. et al., 2021; Pan et al., 2021; Liu J. et al., 2022; Chang et al., 2021; Luo et al., 2022; Wang et al., 2022; Xu et al., 2019; Jiang et al., 2021; Wei et al., 2020; Jiang and Zhang, 2021; Sun and Xu, 2023; Fan et al., 2021; Huang et al., 2021; Patel et al., 2018; Shao et al., 2020; Dong et al., 2022; Yang et al., 2022; Zhao et al., 2020; Xiaoying et al., 2020; Zhao Y. et al., 2022; Liu Y. et al., 2022; Huang W. et al., 2023; Wang Q. S. et al., 2023; Li et al., 2022; Liu B. et al., 2022; Wang Y. et al., 2023; Li et al., 2020; Qian et al., 2022; Shao et al., 2023; Yu et al., 2019; Ji et al., 2019; Zhang and Han, 2022; Shao et al., 2022; Wang et al., 2020) (Table 1).
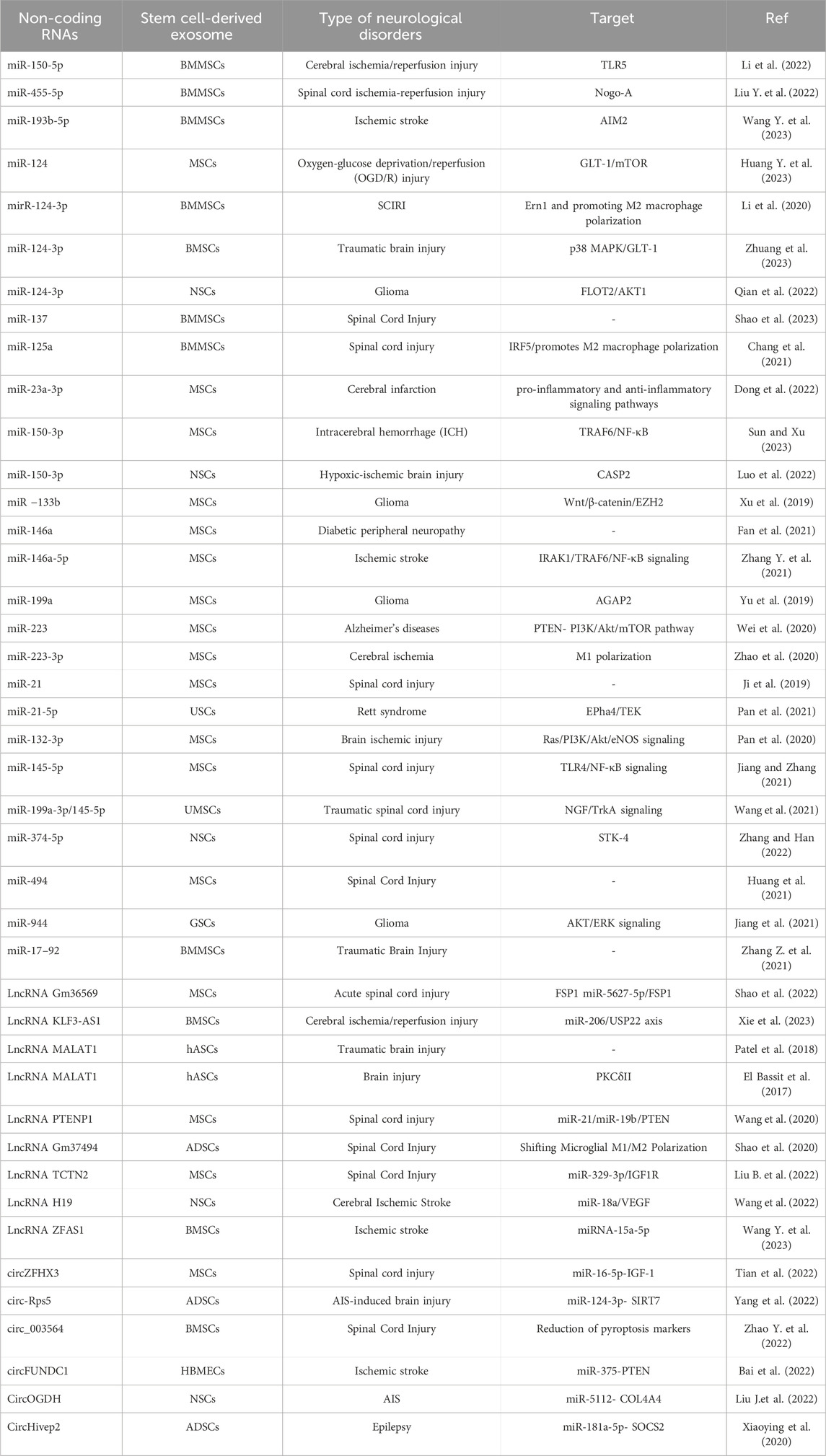
Table 1. Various ncRNAs in different neurological diseases.
6 Alzheimer’s diseaseA disorder known as Alzheimer’s disease (AD) affects the brain and causes deposits of materials known as neurofibrillary tangles and amyloid beta plaques (Scheltens et al., 2021). The possibility of using stem cell-derived exosomes and their contents to help treat AD has been investigated by several researchers. Liu J. et al. (2022) discovered that in an AD model, there was an increase in HIF-1α expression, apoptosis, and a decrease in miR-223. Exosomes generated from MSCs were discovered to be taken up by AD cells, leading to enhanced cell motility, reduced apoptosis, and elevated miR-223 levels. The use of KC7F2, a hypoxia inhibitor, restored these effects. Additional r
Comments (0)