
Remember me
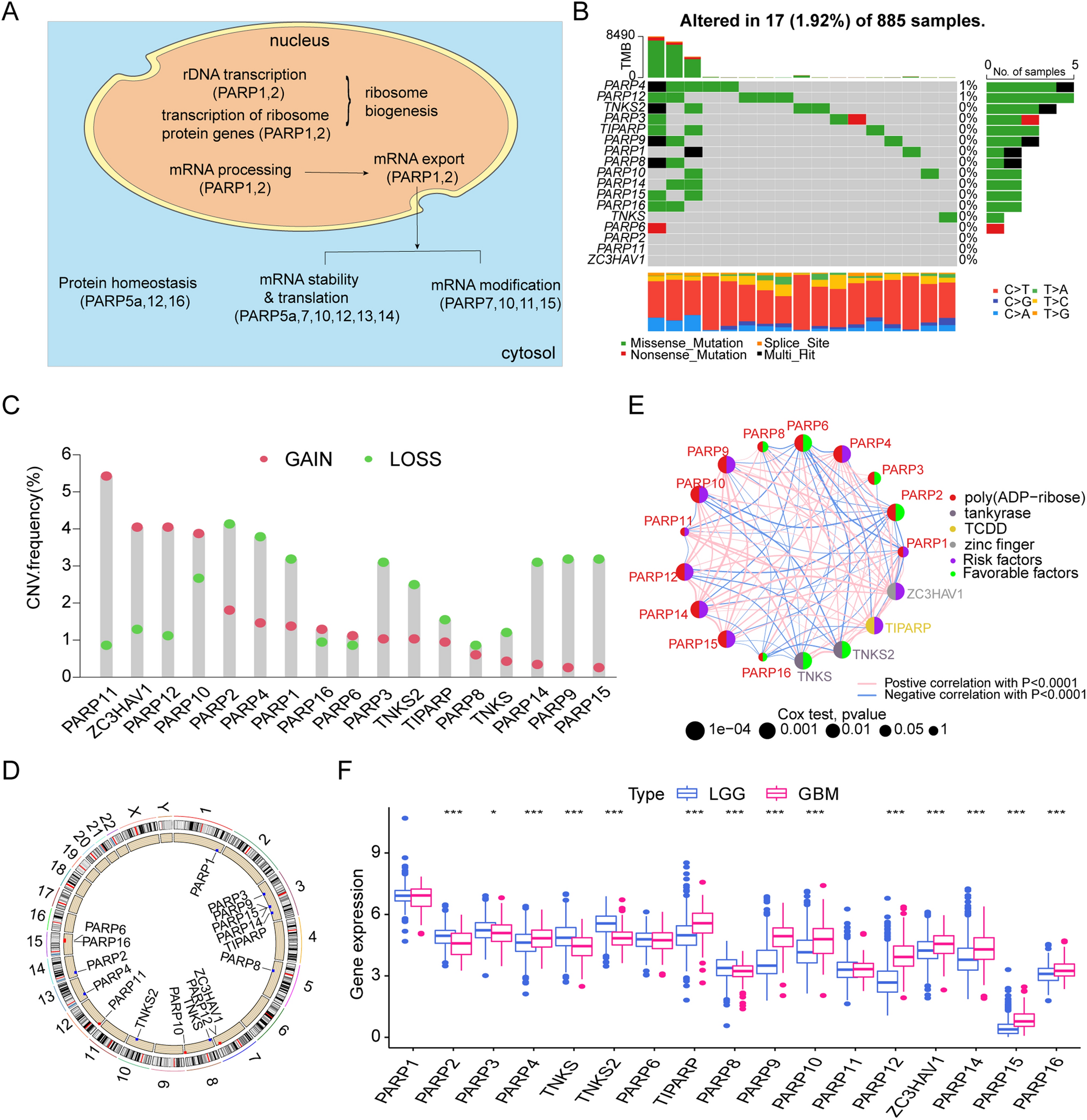

To explore the causal effect of GM on GC, a two-sample MR analysis was performed using IVW and BWMR methods as the main analytical strategies. Supplementary Table 2 contains the precise information on the 4048 SNPs linked to 207 taxa and 205 bacterial pathway traits. As depicted in Fig. 2A, the IVW model identified 25 GMs associated with GC. The BWMR approach tackles the uncertainty surrounding the estimated minor effects and weak horizontal pleiotropic effects, and identifies outliers resulting from significant horizontal pleiotropic effects [23]. Therefore, the IVW results were validated using the BWMR method, which led to the identification of 18 GC-related GMs (Fig. 2B). Finally, a reverse MR study was performed using 18 sets of GM data that were statistically significant to mitigate the potential influence of reverse causation in the context of GC. Supplementary Table 3 shows the SNPs associated with GC. The results of the reverse MR analysis did not indicate of reverse causation (P > 0.05), as shown in Supplementary Table 4.
Fig.2
MR analyses highlight the causal effects between gut microbiota and gastric cancer. a Forest plot of the causal effect of gut microbiota on GC via the IVW method. b Forest plot of the causal effect of gut microbiota on GC via the BWMR method. OR odds ratio, CI confidence interval
As displayed in Fig. 2, MR analysis using the IVW and BWMR methods indicated a genetic prediction of one taxon and three bacterial pathways linked to a higher risk of GC. The presence of g_Haemophilus (IVW OR = 1.4965, 95%CI = 1.0640–2.10480, P = 0.0205; BWMR OR = 1.5121, 95%CI = 1.0537–2.1698, P = 0.0248), “PWY.5659 GDP mannose biosynthesis” (IVW OR = 2.1118, 95%CI = 1.2293–3.6278, P = 0.0068; BWMR OR = 2.1384, 95%CI = 1.2067–3.7897, P = 0.0092), “PWY.724 superpathway of L lysine, L threonine and L methionine biosynthesis II” (IVW OR = 2.3932, 95%CI = 1.5945–1.0623, P = 0.0243; BWMR OR = 1.5768, 95%CI = 1.0472–2.3742, P = 0.0292), and “TRNA CHARGING PWY tRNA charging” (IVW OR = 1.5515, 95%CI = 1.0560–2.2796, P = 0.0253; BWMR OR = 1.5478, 95%CI = 1.0541–2.2729, P = 0.0258) were found to significantly elevate the risk of GC.
Eight taxa and six bacterial pathways were associated with a lower likelihood of developing GC. g_Oxalobacter (IVW OR = 0.7038, 95%CI = 0.5345–0.9269, P = 0.0124; BWMR OR = 0.6964, 95%CI = 0.5204–0.9320, P = 0.0150), s_Alistipes_sp_AP11 (IVW OR = 0.7237, 95%CI = 0.5560–0.9420, P = 0.01620; BWMR OR = 0.7177, 95%CI = 0.5525–0.9322, P = 0.0129), f_Oxalobacteraceae (IVW OR = 0.7040, 95%CI = 0.5255–0.9430, P = 0.0186; BWMR OR = 0.6955, 95%CI = 0.5010–0.9485, P = 0.0218), s_Oxalobacter_formigenes (IVW OR = 0.7039, 95%CI = 0.5253–0.9433, P = 0.0187; BWMR OR = 0.6954, 95%CI = 0.5097–0.948, P = 0.0219), o_Burkholderiales (IVW OR = 0.6640, 95%CI = 0.4615–0.9553, P = 0.0273; BWMR OR = 0.6621, 95%CI = 0.4535–0.9666, P = 0.0327), f_Bacteroidaceae (IVW OR = 0.6483, 95%CI = 0.4380–0.9595, P = 0.0303; BWMR OR = 0.6436, 95%CI = 0.4260–0.9724, P = 0.0364), s_Coprococcus_catus (IVW OR = 0.5983, 95%CI = 0.3690–0.9701, P = 0.0372; BWMR OR = 0.5933, 95%CI = 0.3550–0.9914, P = 0.0463), s_Escherichia_coli (IVW OR = 0.7489, 95%CI = 0.5653–0.9922, P = 0.0439; BWMR OR = 0.7393, 95%CI = 0.5501–0.9937, P = 0.0453), “PWY.6147:6-hydroxymethyl dihydropterin diphosphate biosynthesis I” (IVW OR = 0.5590, 95%CI = 0.3923–0.7966, P = 0.0013; BWMR OR = 0.5719, 95%CI = 0.3973–0.8232, P = 0.0026), “POLYAMSYN.PWY: superpathway of polyamine biosynthesis I” (IVW OR = 0.5003, 95%CI = 0.3118–0.8028, P = 0.0041; BWMR OR = 0.4879, 95%CI = 0.29293 ~ 0.8128, P = 0.0059), “PWY_ARG.POLYAMINE.SYN:superpathway of arginine and polyamine biosynthesis” (IVW OR = 0.4894, 95%CI = 0.2950–0.8120, P = 0.0057; BWMR OR = 0.5000, 95%CI = 0.3025–0.8249, P = 0.0067), “PWY.6897:thiamin salvage II” (IVW OR = 0.5859, 95%CI = 0.3815–0.8996, P = 0.0146; BWMR OR = 0.5770, 95%CI = 0.3661–0.9092, P = 0.0178), “PWY.6284:superpathway of unsaturated fatty acids biosynthesis(E.coli)” (IVW OR = 0.6316, 95%CI = 0.4256–0.9373, P = 0.0225; BWMR OR = 0.6111, 95%CI = 0.4043–0.9237, P = 0.0195), and “ARO.PWY:chorismate biosynthesis I” (IVW OR = 0.6102, 95%CI = 0.3982–0.9352, P = 0.0234; BWMR OR = 0.5961, 95%CI = 0.3757–0.9459, P = 0.0281) were found to significantly decrease in the risk of GC (Fig. 2).
An additional investigation into the causal connection between the 18 GMs and GC is outlined in Fig. 3A–R. Sensitivity analysis provided detailed information confirming the strength of the observed causal relationships. The MR-Egger regression intercept method showed no bias from genetic pleiotropy in the results (Supplementary Table 5), and MR-PRESSO analysis confirmed the lack of horizontal pleiotropy in the MR study (P > 0.05, Supplementary Table 6). The Cochran's Q tests revealed no significant heterogeneity based on the results (P > 0.05; Supplementary Table 7). Furthermore, scatter plots (Fig. S1), forest plots (Fig. S2), and funnel plots (Fig. S3) further supported the stability of the results, and analyses using the “leave-one-out” method indicated that none of the individual variables had a significant impact on the causal inferences (Fig. S4).
Fig.3
Forest plots display the causal associations between gut microbiota and GC using different methods. a Forest plot of the causal effect of the “6-hydroxymethyl dihydropterin diphosphate biosynthesis I” bacterial pathway on GC. b Forest plot of the causal effect of the “superpathway of polyamine biosynthesis I” bacterial pathway on GC. c Forest plot of the causal effect of the “superpathway of arginine and polyamine biosynthesis” bacterial pathway on GC. d Forest plot of the causal effect of the “GDP mannose biosynthesis” bacterial pathway on GC. e Forest plot of the causal effect of the g_Oxalobacter on GC. f Forest plot of the causal effect of the “thiamin salvage II” bacterial pathway on GC. g Forest plot of the causal effect of the s_Alistipes_sp_AP11 on GC. h Forest plot of the causal effect of the f_Oxalobacteraceae on GC. i Forest plot of the causal effect of the s_Oxalobacter_formigenes on GC. j Forest plot of the causal effect of the g_Haemophilus on GC. k Forest plot of the causal effect of the “superpathway of unsaturated fatty acids biosynthesis (E. coli)” bacterial pathway on GC. l Forest plot of the causal effect of the “chorismate biosynthesis I” bacterial pathway on GC. m Forest plot of the causal effect of the “superpathway of L lysine, L threonine and L methionine biosynthesis II” bacterial pathway on GC. n Forest plot of the causal effect of the “TRNA.CHARGING.PWY:tRNA.charging” bacterial pathway on GC. o Forest plot of the causal effect of the o_Burkholderiales on GC. p Forest plot of the causal effect of the f_Bacteroidaceae on GC. q Forest plot of the causal effect of the s_Coprococcus_catus on GC. (r) Forest plot of the causal effect of the s_Escherichia_coli on GC. IVW inverse variance weighting, OR odds ratio, CI confidence interval
3.2 Effect of 18 GMs on 731 immune cell traitsThe crucial role of the immune system in GC is widely recognized. Thus, we proceeded to clarify how the GM affects immune cell characteristics and their potential influence on the risk of GC. Mediation MR analyses were performed with 731 immune cell characteristics as mediators to explore the relationships between the GM and GC. Significant relationships were found between the 18 bacterial traits and various immune cell traits. A total of 467 immune cell characteristics from a pool of 731 potential mediators met the screening criteria (P_IVW < 0.05) and were used in the mediation MR analyses (Supplementary Table 8).
3.3 Effect of each immune cell trait on GCThese results underscore the complex interplay between immune cell characteristics and the GM, laying the groundwork for further mediation analyses. Subsequent investigations revealed the potential mediating effects of exposure to these noteworthy mediators (467 immune cell traits from 18 gut microbiota) in GC. Based on IVW (Fig.S5), 12 GMs affected GC through 16 immune cell traits.
Following the elucidation of how immune cell features affect GC, we examined the direct influence of 12 GM on these crucial mediators (16 immune cell traits). Our analysis revealed several important discoveries (Fig. S6). The causal association between 12 GMs and 16 immune cell traits was investigated using a two-sample MR analysis. The MR-Egger regression intercept method revealed that genetic pleiotropy did not affect these findings (Supplementary Table 9). Moreover, Cochran’s Q tests indicated no significant heterogeneity (P > 0.05, Supplementary Table 10), and MR-PRESSO analysis confirmed the absence of horizontal pleiotropy in the MR study (P > 0.05 Supplementary Table 11). The scatter plots in Fig. S7, forest plots in Fig. S8, funnel plots in Fig. S9, and "leave-one-out" analyses (Fig. S10) highlighted the consistency of the results.
MR analysis was conducted to determine the protective effects of the eight immunophenotypes on GC using the IVW method. These immunophenotypes include (6-hydroxymethyl dihydropterin diphosphate biosynthesis I) CD39 + CD4 + %CD4 + , (f_Oxalobacteraceae) CD4 Treg %T cell, (f_Oxalobacteraceae) CD8br and CD8dim %leukocyte, (s_Oxalobacter_formigenes) CD4 Treg %T cell, (GCST90027810) ebi-a-GCST90001614, (g_Haemophilus) HVEM on naive CD4 + , (chorismate biosynthesis I) CD28- CD8dim %CD8dim, and (tRNA.charging) CD39 + CD4 + %CD4 + . Comparable results were observed when four additional approaches were employed: MR eager, weighted median, simple mode, and weighted mode (Fig. S11).
The IVW approach was employed to examine the enhancing effect of four immunophenotypes on GC, specifically (superpathway of polyamine biosynthesis I) CD8br %T cell, (superpathway of arginine and polyamine biosynthesis) SSC-A on NKT, (thiamin salvage II) CD8br %T cell, (s_Alistipes_sp_AP11) NKT %T cell, (g_Haemophilus) CCR2 on CD14- CD16 + monocyte, (chorismate biosynthesis I) CD28- DN (CD4-CD8-) %T cell, (superpathway of L lysine, L threonine and L methionine biosynthesis II) HLA DR on CD33br HLA DR + CD14dim, and (s_Coprococcus_catus) HLA DR on CD33br HLA DR + CD14dim. Four additional techniques, namely MR eager, weighted median, simple mode, and weighted mode, had similar outcomes (Fig. S11) to the above approaches.
The MR-Egger intercept (Supplementary Table 12) and MR-PRESSO analyses indicated no horizontal pleiotropy in the studied associations (Supplementary Table 13). Cochran's Q tests did not show any significant heterogeneity based on the results shown in Supplementary Table 14 (P > 0.05). The reliability of the findings was reinforced by scatter diagrams (Fig. S12), forest charts (Fig. S13), funnel charts (Fig. S14), and “leave-one-out” examinations (Fig. S15).
3.4 Mediation MR analysisA two-step MR study revealed a causal relationship between the GM and GC, mediated by immune cell traits. The key factors affecting GC were determined, and the resulting impact of exposure on mediation was measured by calculating the mediation effect (indirect effect, Table 1).
Table 1 Mediation Mendelian randomization analyses of the causal effects among gut microbiota, immune cells and GCSome data from Table 1 were briefly explained, particularly “PWY.6147:6-hydroxymethyl dihydropterin diphosphate biosynthesis I” hinders the advancement of GC by interacting with “CD39 + CD4 + %CD4 + ” cells, resulting in a total effect size of -0.5815, of which the mediated effect size of “CD39 + CD4 + %CD4 + ” was 0.0197. Moreover, “POLYAMSYN.PWY: superpathway of polyamine biosynthesis I” was demonstrated to impede the advancement of GC through mediation by “CD8br %T cell”, with a total effect size of -0.6925; the mediated effect size of “CD8br %T cell” was 0.0418. The promotion of GC progression was facilitated by the interaction of “PWY.724: superpathway of L lysine, L threonine and L methionine biosynthesis II” and “HLA DR on CD33br HLA DR + CD14dim”, resulting in a total effect size of 0.4665, with “HLA DR on CD33br HLA DR + CD14dim” cells contributing a mediated effect size of -0.0409. These findings emphasize the intricate relationship between distinct GM exposures, their mediators, and their collective influence on GC.
Comments (0)