
Remember me







To elucidate the molecular mechanisms guiding the adaptive response of colon cancer cells to glucose deprivation, we monitored the gene expression profiles of HCT116 human CRC cells under high- and low-glucose conditions (GSE31084). A substantial elevation in RIOK3 expression was observed under low-glucose conditions compared with that under high-glucose medium (Fig. 1a).
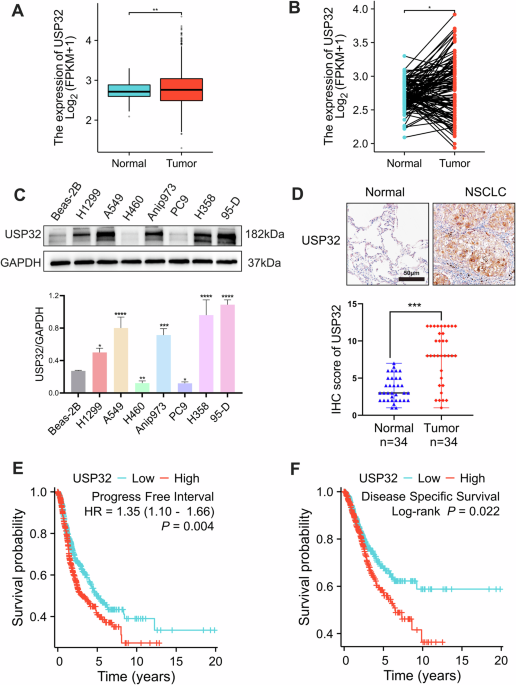
Fig. 1: RIOK3 expression is increased under glucose deficiency.
a RIOK3 mRNA levels increase under low-glucose conditions based on GEO datasets (GSE31084). b HCT116 cells were incubated in low-glucose DMEM for the indicated times. RNA was then extracted and RIOK3 mRNA levels were analyzed with qPCR. Data are shown as mean ± SD (n = 3). c HCT116 cells were incubated in low-glucose DMEM for the indicated times. Western blotting was performed to detect RIOK3 protein levels. d HCT116 cells were cultured in a medium with a specific glucose concentration for 24 h. RNA was then extracted and RIOK3 mRNA levels were analyzed with qPCR. Data are shown as mean ± SD (n = 3). e HCT116 cells were cultured in a medium with a specific glucose concentration for 24 h. Western blotting was performed to detect RIOK3 protein levels.
To validate the impact of glucose deprivation on RIOK3 expression, HCT116 and SW480 colon cancer cells were exposed to a low-glucose (1 g/L) medium. An evident time-dependent increase in RIOK3 mRNA was noted in HCT116 (Fig. 1b). RIOK3 protein levels were also increased in low-glucose medium (Fig. 1c). Consistent results were obtained in SW480 cells, where low-glucose treatment resulted in elevated RIOK3 mRNA and protein levels (Supplementary Fig. S1a, b). We also examined RIOK3 expression in HCT116 cells across varying glucose concentrations, revealing a direct correlation between decreased glucose levels and increased RIOK3 expression in colon cancer cells (Fig. 1d, e).
Additionally, we assessed RIOK3 expression in the normal human colon cell line NCM460 and FHC. As shown in Supplementary Fig. S1c, d, we did not observe significant upregulation of RIOK3 expression upon glucose starvation in either NCM460 or FHC. This suggests that normal cells may employ a unique pathway, distinct from cancer cells, to adapt to glucose starvation.
RIOK3 facilitates cell survival upon glucose deprivationGiven its established role in cancer proliferation and metastasis [6, 7], we investigated whether RIOK3 abundance is imperative for CRC cell survival under glucose limitation.
To this end, we generated RIOK3-KO cell lines using the CRISPR-Cas9 system (Fig. 2a). CCK-8 assay results indicated a significantly higher viability of wild-type (WT) cells compared with that of RIOK3-KO cells under low-glucose conditions (Fig. 2b). We also analyzed the cell death rate under low-glucose conditions and the ability of surviving cells to grow and form colonies. RIOK3-KO HCT116 cells exhibited increased sensitivity to glucose deprivation-induced cell death compared with that in control cells (Fig. 2c and Supplementary Fig. S2a). For the colony formation assay, WT and RIOK3-KO HCT116 cells were subjected to a period of starvation prior to subsequent cultivation in regular growth medium. RIOK3 silencing substantially impaired colony formation in seeded cells under starvation (Fig. 2d), supporting the notion that RIOK3 plays a role in promoting tumor growth and survival, particularly under nutrient-deprived conditions.
Fig. 2: RIOK3 supports cell survival under glucose starvation condition.
a Western blotting was performed to determine RIOK3 protein levels. b WT and RIOK3-KO CRC cells were cultured in low-glucose DMEM for the indicated times. Cell viability was then tested with the CCK-8 assay. Data are shown as mean ± SD (n = 3). c WT and RIOK3-KO HCT116 cells were cultured in low-glucose DMEM for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). d WT and RIOK3-KO HCT116 cells were cultured in high-glucose or low-glucose DMEM, and a clonogenic survival assay was performed. Colonies were counted after crystal violet staining. Right graph, the quantitative data are shown as the mean ± SD (n= 3). e Kaplan–Meier plots were generated to compare the overall survival between the high-expression and low-expression groups of RIOK3.
Furthermore, a prognostic analysis of RIOK3 in colon cancer using the Kaplan –Meier plotter database tool revealed that reduced overall survival in colon cancer patients was linked to the elevated expression level of RIOK3 (Fig. 2e), highlighting the significant role of RIOK3 in CRC progression.
RIOK3 knockout induces ROS generation-triggered cell deathOxidative stress is a primary trigger for cell death under nutrient deficiency [14, 15]. Accordingly, we monitored ROS production following RIOK3 knockout using DCFH-DA fluorescence. Loss of RIOK3 significantly elevated ROS levels in CRC cells, particularly under glucose-deprived conditions (Fig. 3a). Intracellular ROS levels were also assessed at different time points under low glucose conditions, revealing a gradual increase with prolonged exposure to low glucose, closely correlated with RIOK3 expression (Supplementary Fig. S3a). To confirm whether the impact of RIOK3 on cell survival under low-glucose conditions was associated with ROS generation, cells were treated with the ROS scavenger N-acetyl-l-cysteine (NAC), which resulted in the reversal of cell death in RIOK3-KO cells under starvation (Fig. 3b and Supplementary Fig. S3b, c).
Fig. 3: Altered RIOK3 expression affects ROS.
a WT and RIOK3-KO CRC cells were cultured in low-glucose DMEM for 48 h, and the ROS levels were quantified as the mean DCF values. Data are shown as mean ± SD (n = 3). b WT and RIOK3-KO CRC cells were cultured in low-glucose DMEM in the presence or absence of 10 mM NAC for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). c WT and RIOK3-KO CRC cells were cultured in low-glucose DMEM for 48 h, and the NADP+/NADPH ratio was determined. Data are shown as mean ± SD (n = 3). d CRC cells were transfected with an empty vector or flag-RIOK3 plasmids. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h, and the NADP+/NADPH ratio was determined. Data are shown as mean ± SD (n = 3).
NADPH is a key factor in maintaining intracellular redox homeostasis [16]. Therefore, we examined the impact of RIOK3 on intracellular NADPH levels. Indeed, the loss of RIOK3 led to a reduction in NADPH levels, whereas the gain of RIOK3 function consistently elevated them (Fig. 3c, d). This suggests that RIOK3 regulates NADPH homeostasis, thereby enhancing cancer cell survival against oxidative stress.
RIOK3 interacts with HSP90α and subsequently augments IDH1 expressionTo explore the mechanism by which RIOK3 regulates intracellular NADPH levels, we conducted mass spectrometry (MS) analysis to identify its binding partners. MS results revealed an interaction between RIOK3 and HSP90α, a molecular chaperone crucial for various oncogenic processes [17]. Subsequent co-immunoprecipitation (co-IP) experiments were performed to validate the interaction between RIOK3 and HSP90α. Lysates of HCT116 cells expressing flag-RIOK3 were immunoprecipitated with an anti-flag antibody. The immunoprecipitated complex was then analyzed using western blotting with an antibody against HSP90α. As shown in Fig. 4a, these analyses confirmed the physical association between flag-RIOK3 and HSP90α. Employing the RIOK3 antibody for the co-IP assays similarly revealed an observed interaction between RIOK3 and HSP90α (Fig. 4b), providing further evidence of their interplay in the cellular context.
Fig. 4: High expression of RIOK3 promotes the binding between HSP90α and IDH1.
a HCT116 cells were transfected with an empty vector or flag-RIOK3 plasmids. 24 h after transfection, the cell lysate was extracted for co-IP with anti-flag antibody. b HCT116 cell lysates were extracted for co-IP using anti-RIOK3 antibody. c HCT116 cell lysates were extracted for co-IP using anti-HSP90α antibody. d HCT116 cells were transfected with an empty vector or flag-RIOK3 plasmids. 24 h after transfection, the cell lysate was extracted for co-IP with anti-HSP90α antibody. e WT and RIOK3-KO HCT116 cells were cultured in low-glucose DMEM for 48 h, and the cell lysate was extracted for co-IP with anti-HSP90α antibody. f WT and RIOK3-KO HCT116 cells were cultured in low-glucose DMEM for 48 h, western blotting was performed to determine IDH1, HSP90α, and RIOK3 protein levels. g WT and RIOK3-KO HCT116 cells were treated with or without 50 μM CHX, and western blotting was used to detect IDH1 level. h WT and RIOK3-KO HCT116 cells were cultured in low-glucose DMEM for 48 h and treated with or without 2 μM MG132, and western blotting was used to detect IDH1 level. i. WT and RIOK3-KO HCT116 cells expressing HA-Ubiquitin plasmids were cultured in low-glucose DMEM for 48 h with 2 μM MG132, and the cell lysate was extracted for co-IP with anti-IDH1 antibody. j Western blotting was performed to determine HSP90α protein levels. k HCT116 cells expressing control-shRNA or HSP90α-shRNA were cultured in low-glucose DMEM for 48 h, and western blotting was performed to determine IDH1 and HSP90α protein levels. m HCT116 cells expressing control-shRNA or HSP90α-shRNA were transfected with a non-specific siRNA or RIOK3 siRNA. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h. Western blotting was performed to determine IDH1, HSP90α, and RIOK3 protein levels.
The NADPH-producing enzyme IDH1 is reportedly an interacting partner of HSP90 [18]. Therefore, we validated the interaction between HSP90α and IDH1 in colon cancer cells. The co-IP experiment demonstrated the binding of HSP90α to IDH1 (Fig. 4c). Notably, RIOK3 overexpression enhanced both IDH1 protein levels and its interaction with HSP90α (Fig. 4d). Subsequently, we examined the binding between HSP90α and IDH1 in RIOK3-KO HCT116 cells. As can be seen in Fig. 4e, an interaction between IDH1 and HSP90α was observed in a low-glucose environment. However, in RIOK3-KO cells, both the expression of IDH1 and its binding with HSP90α were substantially decreased. Therefore, we hypothesized that RIOK3 modulates IDH1 expression by influencing its interaction with HSP90α in response to glucose availability.
Concurrently evaluating the influence of different glucose concentrations on IDH1 expression, we observed an increase in IDH1 expression as glucose concentration decreased, consistent with RIOK3 (Supplementary Fig. S4a). To further validate this regulatory role of RIOK3, we treated the RIOK3-KO cell line with low-glucose culture medium and confirmed that low glucose conditions failed to induce the expression of IDH1 in RIOK3-KO cells (Fig. 4f). We hypothesized that RIOK3 might influence the expression of IDH1 by affecting its stability. To validate this hypothesis, we conducted a cycloheximide (CHX) chase experiment and observed a significantly accelerated degradation rate of IDH1 in RIOK3-KO cells (Fig. 4g), indicating that RIOK3 exerts an impact on IDH1 expression by affecting its stability. To further clarify the mechanism through which RIOK3 regulates IDH1 stability and considering potential ubiquitination sites on IDH1 [19], we treated HCT116 cells with MG132, a cell-permeable proteasome inhibitor, to determine whether IDH1 could undergo ubiquitination degradation. As illustrated in Fig. 4h, MG132 restored reduced IDH1 levels caused by RIOK3-KO under low-glucose conditions. Subsequently, we transfected HA-ubiquitin plasmids into both WT and RIOK3-KO HCT116 cells and observed increased ubiquitination of IDH1 in RIOK3-KO cells (Fig. 4i). This indicates that RIOK3 enhances IDH1 stability by reducing its ubiquitination levels.
To further investigate the role of HSP90α, HSP90α-knockdown (KD) HCT116 cell lines were generated using shRNA (Fig. 4j and Supplementary Fig. S4b). Intriguingly, we observed a decrease in IDH1 expression in HSP90α-KD cells (Fig. 4k), yet its expression was unaffected by RIOK3 (Fig. 4m). We further validated the impact of HSP90 activity on IDH1 expression using the HSP90 inhibitor 17-AAG and observed that 17-AAG treatment suppressed IDH1 expression, and low glucose did not lead to a significant increase in IDH1 expression (Supplementary Fig. S4c). Furthermore, 17-AAG treatment attenuated the differences in IDH1 expression between WT and RIOK3-KO cells (Supplementary Fig. S4d), further confirming that RIOK3 modulates IDH1 expression through HSP90α.
RIOK3 regulates NADPH production and maintains redox balance through HSP90αTo further elucidate whether the impact of RIOK3 on NADPH production is linked to its binding to HSP90α, we generated IDH1-KD HCT116 cell lines using shRNA (Fig. 5a and Supplementary Fig. S5a). Knocking down either HSP90α or IDH1 decreased NADPH levels in colon cancer cells, especially under low-glucose conditions (Fig. 5b). RIOK3 inhibition did not significantly affect NADPH levels in HSP90α-KD colon cancer cells compared with that in the control cells (Fig. 5c), suggesting that HSP90α plays a crucial role in mediating RIOK3’s impact on NADPH metabolism. Concordantly, the utilization of 17-AAG similarly attenuated the impact of RIOK3 on NADPH (Supplementary Fig. S5b). Similarly, RIOK3 inhibition failed to induce a substantial disparity in NADPH levels in IDH1-KD CRC cells compared with that in the control group (Fig. 5d). Subsequently, we overexpressed flag-RIOK3 in HSP90α-KD and IDH1-KD cells and observed that overexpression of RIOK3 in low-glucose environments led to increased NADPH levels, and this effect was attenuated when HSP90α or IDH1 was knocked down (Fig. 5e). Furthermore, IDH1 overexpression in RIOK3-KO cells partially restored the inhibitory effect of RIOK3 on NADPH (Fig. 5f), further supporting the notion that the regulatory effect of RIOK3 on NADPH production likely depends on HSP90α and IDH1.
Fig. 5: RIOK3 modulates NADPH generation and preserves redox equilibrium via HSP90α.
a Western blotting was performed to determine IDH1 protein levels. b HCT116 cells expressing control-shRNA, HSP90α-shRNA, or IDH1-shRNA were cultured in low-glucose DMEM for 48 h, and NADP+/NADPH ratio was determined. Data are shown as mean ± SD (n = 3). HCT116 cells expressing control-shRNA, HSP90α-shRNA (c), or IDH1-shRNA (d) were transfected with a non-specific siRNA or RIOK3 siRNA. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h. The NADP+/NADPH ratio was then determined. Data are shown as mean ± SD (n = 3). e HCT116 cells expressing control-shRNA, HSP90α-shRNA, or IDH1-shRNA were transfected with an empty vector or flag-RIOK3 plasmids. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h. The NADP+/NADPH ratio was then determined. Data are shown as mean ± SD (n = 3). f WT and RIOK3-KO HCT116 cells were transfected with an empty vector or flag-IDH1 plasmids, 24 h after transfection, and cells were cultured in low-glucose DMEM for 48 h. The NADP+/NADPH ratio was then determined. Data are shown as mean ± SD (n = 3). g HCT116 cells expressing control-shRNA, HSP90α-shRNA, or IDH1-shRNA were cultured in low-glucose DMEM for 48 h, and the ROS levels were quantified as the mean DCF values. Data are shown as mean ± SD (n = 3). HCT116 cells expressing control-shRNA, HSP90α-shRNA (h), or IDH1-shRNA (i) were transfected with a non-specific siRNA or RIOK3 siRNA. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h. ROS levels were then quantified as the mean DCF values. Data are shown as mean ± SD (n = 3). j HCT116 cells expressing control-shRNA, HSP90α-shRNA, or IDH1-shRNA were transfected with an empty vector or flag-RIOK3 plasmids. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h. ROS levels were then quantified as the mean DCF values. Data are shown as mean ± SD (n = 3). k WT and RIOK3-KO HCT116 cells were transfected with an empty vector or flag-IDH1 plasmids, 24 h after transfection, and cells were cultured in low-glucose DMEM for 48 h. ROS levels were then quantified as the mean DCF values. Data are shown as mean ± SD (n = 3).
We also observed that depletion of either HSP90α or IDH1 led to differential ROS elevation in HSP90α-KD and IDH1-KD cells, respectively (Fig. 5g). Notably, RIOK3 inhibition did significantly alter the ROS levels in these cells compared with that in control cells (Fig. 5h, i). We observed comparable ROS levels in WT and RIOK3-KO cells post-17-AAG treatment (Supplementary Fig. S5c). The influence of RIOK3 overexpression on ROS production was mitigated by HSP90α or IDH1 knockdown (Fig. 5j). Notably, IDH1 overexpression in RIOK3-KO cells partially reversed RIOK3’s inhibitory influence on ROS (Fig. 5k).
Collectively, these findings imply that RIOK3 plays a pivotal role in mediating the maintenance of redox balance and regulation of ROS levels in colon cancer cells through HSP90α- and IDH1-dependent mechanisms.
RIOK3 supports colon cancer cell survival under glucose starvation via HSP90αAn increased cell death rate in HSP90α-KD cells under low-glucose conditions compared with that in the control group was observed (Fig. 6a and Supplementary Fig. S6a). Interestingly, RIOK3 knockdown did not significantly impact cell death rates in HSP90α-KD cells (Fig. 6b and Supplementary Fig. S6b). Likewise, treating RIOK3-KO cells with the HSP90 inhibitor 17-AAG diminished the differences in cell death rates (Fig. 6c and Supplementary Fig. S6c), suggesting that RIOK3 inhibition did not substantially impact cell death in the absence of HSP90. Similarly, cell death increased in IDH1-KD cells compared with that in the control cells (Fig. 6d and Supplementary Fig. S6d). However, we observed that cell death rates did not significantly differ in IDH1-KD cells with either negative control or RIOK3 knockdown, indicating that RIOK3 knockdown did not significantly affect cell death in the absence of IDH1 (Fig. 6e and Supplementary Fig. S6e). Furthermore, IDH1 overexpression in RIOK3-KO HCT116 cells partially counteracted the inhibitory effects of RIOK3 knockout on the viability of colon cancer cells (Fig. 6f and Supplementary Fig. S6f).
Fig. 6: RIOK3‐promoted cell survival under glucose starvation is mediated via HSP90α.
a HCT116 cells expressing control-shRNA or HSP90α-shRNA were cultured in low-glucose DMEM for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). b HCT116 cells expressing control-shRNA or HSP90α-shRNA were transfected with a non-specific siRNA or RIOK3 siRNA. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). c WT and RIOK3-KO HCT116 cells were cultured in low-glucose DMEM with 400 nM 17-AAG for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). d HCT116 cells expressing control-shRNA or IDH1-shRNA were cultured in low-glucose DMEM for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). e HCT116 cells expressing control-shRNA or IDH1-shRNA were transfected with a non-specific siRNA or RIOK3 siRNA. 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). f WT and RIOK3-KO HCT116 cells were transfected with an empty vector or flag-IDH1 plasmids, 24 h after transfection, cells were cultured in low-glucose DMEM for 48 h, and the cell death rate was measured by 7-AAD staining assay. Data are shown as mean ± SD (n = 3). g WT and RIOK3-KO HCT116 cells were injected into BALB/c nude mice, and tumor growth was monitored (n = 4). h, i Xenograft weight and size were measured (n = 4). j Protein was extracted from the tumors and analyzed with western blotting. k The tumor tissues were harvested for NADP+/NADPH detection. Data are shown as mean ± SD (n = 4).
We validated the effect of RIOK3 on colon cancer development in vivo using a nude mouse tumorigenesis experiment. Loss of RIOK3 inhibits tumor growth, whereas intraperitoneal injection of HSP90 inhibitor 17-AAG diminished the disparity in tumor size between WT and RIOK3-KO cells (Fig. 6g–i)). IDH1 expression and intratumoral NADPH levels in subcutaneous tumors of nude mice were assessed simultaneously. Consistent with the findings from in vitro cell experiments, RIOK3-KO tumors exhibited reduced IDH1 expression (Fig. 6j), accompanied by a decrease in NADPH levels (Fig. 6k). Furthermore, immunohistochemistry experiments to assess RIOK3 and IDH1 expression in WT tumors (Supplementary Fig. S7a) revealed increased expression levels of both RIOK3 and IDH1 in the central region of the tumor, consistent with the presence of a glucose-deficient environment in this region (Supplementary Fig. S7b).
In summary, our findings revealed that the role of RIOK3 in enhancing the tolerance of colon cancer cells to glucose deprivation is contingent on HSP90α activity.
Comments (0)